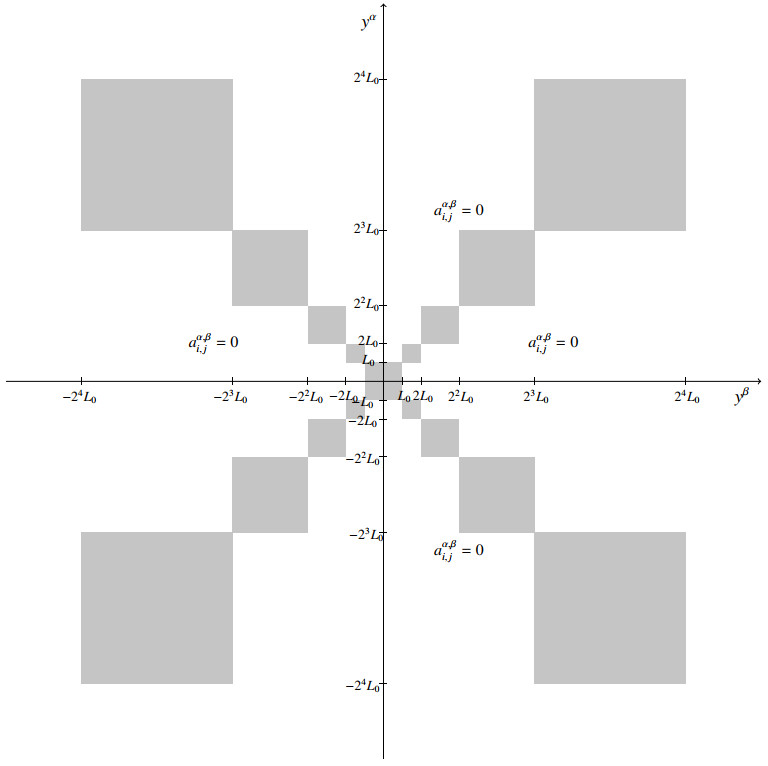
We prove the existence of a solution to a quasilinear system of degenerate equations, when the datum is in a Marcinkiewicz space. The main assumption asks the off-diagonal coefficients to have support in the union of a geometric progression of squares.
Citation: Patrizia Di Gironimo, Salvatore Leonardi, Francesco Leonetti, Marta Macrì, Pier Vincenzo Petricca. Existence of solutions to some quasilinear degenerate elliptic systems with right hand side in a Marcinkiewicz space[J]. Mathematics in Engineering, 2023, 5(3): 1-23. doi: 10.3934/mine.2023055
We prove the existence of a solution to a quasilinear system of degenerate equations, when the datum is in a Marcinkiewicz space. The main assumption asks the off-diagonal coefficients to have support in the union of a geometric progression of squares.

| [1] | S. Aouaoui, Solutions to quasilinear equations of $N$-biharmonic type with degenerate coercivity, Electronic Journal of Differential Equations, 2014 (2014), 228. |
| [2] |
A. Alvino, L. Boccardo, V. Ferone, L. Orsina, G. Trombetti, Existence results for nonlinear elliptic equations with degenerate coercivity, Annali di Matematica, 182 (2003), 53–79. https://doi.org/10.1007/s10231-002-0056-y doi: 10.1007/s10231-002-0056-y

|
| [3] | A. Alvino, V. Ferone, G. Trombetti, A priori estimates for a class of non uniformly elliptic equations, Atti Sem. Mat. Fis. Univ. Modena, 46 (1998), 381–391. |
| [4] | H. Ayadi, F. Mokhtari, Nonlinear anisotropic elliptic equations with variable exponents and degenerate coercivity, Electronic Journal of Differential Equations, 2018 (2018), 45. |
| [5] |
P. Baroni, M. Colombo, G. Mingione, Regularity for general functionals with double phase, Calc. Var., 57 (2018), 62. https://doi.org/10.1007/s00526-018-1332-z doi: 10.1007/s00526-018-1332-z
|
| [6] |
L. Beck, G. Mingione, Lipschitz bounds and nonuniform ellipticity, Commun. Pure Appl. Math., 73 (2020), 944–1034, https://doi.org/10.1002/cpa.21880 doi: 10.1002/cpa.21880
|
| [7] | L. Boccardo, Quasilinear elliptic equations with natural growth terms: the regularizing effect of the lower order terms, J. Nonlinear Convex Anal., 7 (2006), 355–365. |
| [8] |
L. Boccardo, Marcinkiewicz estimates for solutions of some elliptic problems with nonregular data, Annali di Matematica, 188 (2009), 591–601. https://doi.org/10.1007/s10231-008-0090-5 doi: 10.1007/s10231-008-0090-5
|
| [9] | L. Boccardo, H. Brézis, Some remarks on a class of elliptic equations with degenerate coercivity, Boll. Unione Mat. Ital., 6 (2003), 521–530. |
| [10] | L. Boccardo, G. Croce, Elliptic partial differential equations: Existence and regularity of distributional solutions, Berlin: De Gruyter, 2013. https://doi.org/10.1515/9783110315424 |
| [11] |
L. Boccardo, G. Croce, L. Orsina, Existence of solutions for some noncoercive elliptic problems involving derivatives of nonlinear terms, Differential Equations & Applications, 4 (2012), 3–9. https://doi.org/10.7153/dea-04-02 doi: 10.7153/dea-04-02
|
| [12] |
L. Boccardo, G. Croce, L. Orsina, Nonlinear degenerate elliptic problems with $W^{1, 1}_0(\Omega)$ solutions, Manuscripta Math., 137 (2012), 419–439. https://doi.org/10.1007/s00229-011-0473-6 doi: 10.1007/s00229-011-0473-6
|
| [13] | L. Boccardo, G. Croce, C. Tanteri, An elliptic system with degenerate coercivity, Rend. Mat. Appl. (7), 36 (2015), 1–9. |
| [14] | L. Boccardo, A. Dall'Aglio, L. Orsina, Existence and regularity results for some elliptic equations with degenerate coercivity, Atti Sem. Mat. Fis. Univ. Modena, 46 (1998), 51–81. |
| [15] |
P. Cianci, G. R. Cirmi, S. D'Asero, S. Leonardi, Morrey estimates for solutions of singular quadratic nonlinear equations, Annali di Matematica, 196 (2017), 1739–1758. https://doi.org/10.1007/s10231-017-0636-5 doi: 10.1007/s10231-017-0636-5
|
| [16] |
G. R. Cirmi, S. D'Asero, S. Leonardi, Fourth-order nonlinear elliptic equations with lower order term and natural growth conditions, Nonlinear Anal. Theor., 108 (2014), 66–86. https://doi.org/10.1016/j.na.2014.05.014 doi: 10.1016/j.na.2014.05.014
|
| [17] |
G. R. Cirmi, S. D'Asero, S. Leonardi, Morrey estimates for a class of elliptic equations with drift term, Adv. Nonlinear Anal., 9 (2020), 1333–1350. https://doi.org/10.1515/anona-2020-0055 doi: 10.1515/anona-2020-0055
|
| [18] |
G. R. Cirmi, S. D'Asero, S. Leonardi, On the existence of weak solutions to a class of nonlinear elliptic systems with drift term, J. Math. Anal. Appl., 491 (2020), 124370. https://doi.org/10.1016/j.jmaa.2020.124370 doi: 10.1016/j.jmaa.2020.124370
|
| [19] |
G. R. Cirmi, S. D'Asero, S. Leonardi, Morrey estimates for a class of noncoercive elliptic systems with VMO-coefficients, Atti Accad. Naz. Lincei Rend. Lincei Mat. Appl., 32 (2021), 317–334. https://doi.org/10.4171/RLM/938 doi: 10.4171/RLM/938
|
| [20] |
G. R. Cirmi, S. D'Asero, S. Leonardi, M. M. Porzio, Local regularity results for solutions of linear elliptic equations with drift term, Adv. Calc. Var., 15 (2022), 19–32. https://doi.org/10.1515/acv-2019-0048 doi: 10.1515/acv-2019-0048
|
| [21] |
G. R. Cirmi, S. Leonardi, Regularity results for the gradient of solutions linear elliptic equations with $L^{1, \lambda}$ data, Annali di Matematica, 185 (2006), 537–553. http://doi.org/10.1007/s10231-005-0167-3 doi: 10.1007/s10231-005-0167-3
|
| [22] |
G. R. Cirmi, S. Leonardi, Regularity results for solutions of nonlinear elliptic equations with $L^{1, \lambda}$ data, Nonlinear Anal. Theor., 69 (2008), 230–244. https://doi.org/10.1016/j.na.2007.05.014 doi: 10.1016/j.na.2007.05.014
|
| [23] |
G. R. Cirmi, S. Leonardi, Higher differentiability for the solutions of nonlinear elliptic systems with lower-order terms and $L^{1, \theta}$-data, Annali di Matematica, 193 (2014), 115–131. https://doi.org/10.1007/s10231-012-0269-7 doi: 10.1007/s10231-012-0269-7
|
| [24] |
G. Croce, An elliptic problem with degenerate coercivity and a singular quadratic gradient lower order term, Discrete Contin. Dyn. Syst. S, 5 (2012), 507–530. https://doi.org/10.3934/dcdss.2012.5.507 doi: 10.3934/dcdss.2012.5.507
|
| [25] | G. Croce, The regularizing effects of some lower order terms in an elliptic equation with degenerate coercivity, Rendiconti di Matematica, Serie Ⅶ, 27 (2007), 299–314. |
| [26] |
G. Cupini, F. Giannetti, R. Giova, A. Passarelli di Napoli, Regularity results for vectorial minimizers of a class of degenerate convex integrals, J. Differ. Equations, 265 (2018), 4375–4416. https://doi.org/10.1016/j.jde.2018.06.010 doi: 10.1016/j.jde.2018.06.010
|
| [27] |
G. Cupini, P. Marcellini, E. Mascolo, Regularity of minimizers under limit growth conditions, Nonlinear Anal. Theor., 153 (2017), 294–310. https://doi.org/10.1016/j.na.2016.06.002 doi: 10.1016/j.na.2016.06.002
|
| [28] |
C. De Filippis, G. Mingione, Lipschitz bounds and nonautonomous integrals, Arch. Rational Mech. Anal., 242 (2021), 973–1057. https://doi.org/10.1007/s00205-021-01698-5 doi: 10.1007/s00205-021-01698-5
|
| [29] | E. De Giorgi, Un esempio di estremali discontinue per un problema variazionale di tipo ellittico, Boll. Un. Mat. Ital. Ser. Ⅳ, 1 (1968), 135–137. |
| [30] |
F. Della Pietra, G. Di Blasio, Comparison, existence and regularity results for a class of non-uniformly elliptic equations, Differential Equations & Applications, 2 (2010), 79–103. https://doi.org/10.7153/dea-02-07 doi: 10.7153/dea-02-07
|
| [31] | P. Di Gironimo, F. Leonetti, M. Macrì, P. V. Petricca, Existence of bounded solutions for some quasilinear degenerate elliptic systems, Minimax Theory and its Applications, 6 (2021), 321–340. |
| [32] | P. Di Gironimo, F. Leonetti, M. Macrì, P. V. Petricca, Existence of solutions to some quasilinear degenerate elliptic systems when the datum has an intermediate degree of integrability, Complex Var. Elliptic Equ., in press. https://doi.org/10.1080/17476933.2022.2069753 |
| [33] |
G. Dolzmann, N. Hungerbuhler, S. Muller, Non-linear elliptic systems with measure-valued right hand side, Math. Z., 226 (1997), 545–574. https://doi.org/10.1007/PL00004354 doi: 10.1007/PL00004354
|
| [34] |
V. Ferone, N. Fusco, VMO solutions of the $N$-Laplacian with measure data, Comptes Rendus de l'Académie des Sciences Series I-Mathematics, 325 (1997), 365–370. https://doi.org/10.1016/S0764-4442(97)85618-2 doi: 10.1016/S0764-4442(97)85618-2
|
| [35] |
H. Gao, M. Huang, W. Ren, Regularity for entropy solutions to degenerate elliptic equations, J. Math. Anal. Appl., 491 (2020), 124251. https://doi.org/10.1016/j.jmaa.2020.124251 doi: 10.1016/j.jmaa.2020.124251
|
| [36] |
H. Gao, F. Leonetti, W. Ren, Regularity for anisotropic elliptic equations with degenerate coercivity, Nonlinear Anal., 187 (2019), 493–505. https://doi.org/10.1016/j.na.2019.06.017 doi: 10.1016/j.na.2019.06.017
|
| [37] |
D. Giachetti, M. M. Porzio, Existence results for some nonuniformly elliptic equations with irregular data, J. Math. Anal. Appl., 257 (2001), 100–130. https://doi.org/10.1006/jmaa.2000.7324 doi: 10.1006/jmaa.2000.7324
|
| [38] |
D. Giachetti, M. M. Porzio, Elliptic equations with degenerate coercivity: gradient regularity, Acta Math. Sinica, 19 (2003), 349–370. https://doi.org/10.1007/s10114-002-0235-1 doi: 10.1007/s10114-002-0235-1
|
| [39] | W. Hao, S. Leonardi, J. Nečas, An example of irregular solution to a nonlinear Euler-Lagrange elliptic system with real analytic coefficients, Annali della Scuola Normale Superiore di Pisa (Ⅳ), 23 (1996), 57–67. |
| [40] | W. Hao, S. Leonardi, M. Steinhauer, Examples of discontinuous, divergence-free solutions to elliptic variational problems, Comment. Math. Univ. Carolin., 36 (1995), 511–517. |
| [41] |
J. Kristensen, G. Mingione, The singular set of $\omega$-minima, Arch. Rational Mech. Anal., 177 (2005), 93–114. https://doi.org/10.1007/s00205-005-0361-x doi: 10.1007/s00205-005-0361-x
|
| [42] |
J. Kristensen, G. Mingione, The singular set of minima of integral functionals, Arch. Rational Mech. Anal., 180 (2006), 331–398. https://doi.org/10.1007/s00205-005-0402-5 doi: 10.1007/s00205-005-0402-5
|
| [43] |
J. Kristensen, G. Mingione, Boundary regularity in variational problems, Arch. Rational Mech. Anal., 198 (2010), 369–455. https://doi.org/10.1007/s00205-010-0294-x doi: 10.1007/s00205-010-0294-x
|
| [44] |
T. Kuusi, G. Mingione, Universal potential estimates, J. Funct. Anal., 262 (2012), 4205–4269. https://doi.org/10.1016/j.jfa.2012.02.018 doi: 10.1016/j.jfa.2012.02.018
|
| [45] |
T. Kuusi, G. Mingione, Linear potentials in nonlinear potential theory, Arch. Rational Mech. Anal., 207 (2013), 215–246. https://doi.org/10.1007/s00205-012-0562-z doi: 10.1007/s00205-012-0562-z
|
| [46] |
T. Kuusi, G. Mingione, Guide to nonlinear potential estimates, Bull. Math. Sci., 4 (2014), 1–82. https://doi.org/10.1007/s13373-013-0048-9 doi: 10.1007/s13373-013-0048-9
|
| [47] |
T. Kuusi, G. Mingione, The Wolff gradient bound for degenerate parabolic equations, J. Eur. Math. Soc., 16 (2014), 835–892. https://doi.org/10.4171/JEMS/449 doi: 10.4171/JEMS/449
|
| [48] |
T. Kuusi, G. Mingione, Vectorial nonlinear potential theory, J. Eur. Math. Soc., 20 (2018), 929–1004. https://doi.org/10.4171/JEMS/780 doi: 10.4171/JEMS/780
|
| [49] |
S. Leonardi, On constants of some regularity theorems. De Giorgi's type counterexample, Math. Nachr., 192 (1998), 191–204. https://doi.org/10.1002/mana.19981920111 doi: 10.1002/mana.19981920111
|
| [50] |
S. Leonardi, Gradient estimates below duality exponent for a class of linear elliptic systems, Nonlinear Differ. Equ. Appl., 18 (2011), 237–254. http://doi.org/10.1007/s00030-010-0093-y doi: 10.1007/s00030-010-0093-y
|
| [51] |
S. Leonardi, Morrey estimates for some classes of elliptic equations with a lower order term, Nonlinear Anal., 177 (2018), 611–627. https://doi.org/10.1016/j.na.2018.05.010 doi: 10.1016/j.na.2018.05.010
|
| [52] |
S. Leonardi, F. Leonetti, C. Pignotti, E. Rocha, V. Staicu, Maximum principles for some quasilinear elliptic systems, Nonlinear Anal., 194 (2020), 111377. https://doi.org/10.1016/j.na.2018.11.004 doi: 10.1016/j.na.2018.11.004
|
| [53] | S. Leonardi, F. Leonetti, C. Pignotti, E. Rocha, V. Staicu, Local boundedness for weak solutions to some quasilinear elliptic systems, Minimax Theory and its Applications, 6 (2021), 365–378. |
| [54] |
S. Leonardi, F. Leonetti, E. Rocha, V. Staicu, Butterfly support for off diagonal coefficients and boundedness of solutions to quasilinear elliptic systems, Adv. Nonlinear Anal., 11 (2022), 672–683. https://doi.org/10.1515/anona-2021-0205 doi: 10.1515/anona-2021-0205
|
| [55] |
F. Leonetti, P. V. Petricca, Existence of bounded solutions to some nonlinear degenerate elliptic systems, Discrete Contin. Dyn. Syst. B, 11 (2009), 191–203. https://doi.org/10.3934/dcdsb.2009.11.191 doi: 10.3934/dcdsb.2009.11.191
|
| [56] |
F. Leonetti, E. Rocha, V. Staicu, Quasilinear elliptic systems with measure data, Nonlinear Anal. Theor., 154 (2017), 210–224. https://doi.org/10.1016/j.na.2016.04.002 doi: 10.1016/j.na.2016.04.002
|
| [57] |
F. Leonetti, E. Rocha, V. Staicu, Smallness and cancellation in some elliptic systems with measure data, J. Math. Anal. Appl., 465 (2018), 885–902. https://doi.org/10.1016/j.jmaa.2018.05.047 doi: 10.1016/j.jmaa.2018.05.047
|
| [58] |
F. Leonetti, R. Schianchi, A remark on some degenerate elliptic problems, Ann. Univ. Ferrara, 44 (1998), 123–128. https://doi.org/10.1007/BF02828019 doi: 10.1007/BF02828019
|
| [59] |
J. Leray, J. L. Lions, Quelques résultats de Višik sur les problèmes elliptiques non linéaires par les méthodes de Minty-Browder, Bulletin de la Société Mathématique de France, 93 (1965), 97–107. https://doi.org/10.24033/bsmf.1617 doi: 10.24033/bsmf.1617
|
| [60] |
V. G. Maz'ja, Examples of nonregular solutions of quasilinear elliptic equations with analytic coefficients, Funct. Anal. Appl., 2 (1968), 230–234. https://doi.org/10.1007/BF01076124 doi: 10.1007/BF01076124
|
| [61] |
G. Mingione, Regularity of minima: An invitation to the dark side of the calculus of variations, Appl. Math., 51 (2006), 355–426. https://doi.org/10.1007/s10778-006-0110-3 doi: 10.1007/s10778-006-0110-3
|
| [62] |
G. Mingione, The Calder$\acute{o}$n-Zygmund theory for elliptic problems with measure data, Ann. Scuola Norm. Sup. Pisa Cl. Sci. (5), 6 (2007), 195–261. https://doi.org/10.2422/2036-2145.2007.2.01 doi: 10.2422/2036-2145.2007.2.01
|
| [63] |
G. Mingione, Gradient estimates below the duality exponent, Math. Ann., 346 (2010), 571–627. https://doi.org/10.1007/s00208-009-0411-z doi: 10.1007/s00208-009-0411-z
|
| [64] |
G. Mingione, Gradient potential estimates, J. Eur. Math. Soc., 13 (2011), 459–486. https://doi.org/10.4171/JEMS/258 doi: 10.4171/JEMS/258
|
| [65] |
G. Mingione, G. Palatucci, Developments and perspectives in nonlinear potential theory, Nonlinear Anal., 194 (2020), 111452. https://doi.org/10.1016/j.na.2019.02.006 doi: 10.1016/j.na.2019.02.006
|
| [66] | J. Nečas, J. Stará, Principio di massimo per i sistemi ellittici quasi-lineari non diagonali, Boll. Un. Mat. Ital. Ser. Ⅳ, 6 (1972), 1–10. |
| [67] | A. Porretta, Uniqueness and homogenization for a class of non coercive operators in divergence form, Atti Sem. Mat. Fis. Univ. Modena, 46 (1998), 915–936. |
| [68] | E. Stein, G. Weiss, Introduction to Fourier analysis on Euclidean spaces, Princeton University Press, 1971. |
| [69] | C. Trombetti, Existence and regularity for a class of non-uniformly elliptic equations in two dimensions, Differential Integral Equations, 13 (2000), 687–706. |
| [70] | Z. Q. Yan, Everywhere regularity for solutions to quasilinear elliptic systems of triangular form, In: Partial differential equations, Berlin: Springer, 1988,255–261. https://doi.org/10.1007/BFb0082938 |
| [71] | S. Zhou, A note on nonlinear elliptic systems involving measures, Electronic Journal of Differential Equations, 2000 (2000), 08. |
Figures(1)

Patrizia Di Gironimo, Salvatore Leonardi, Francesco Leonetti, Marta Macrì, Pier Vincenzo Petricca. Existence of solutions to some quasilinear degenerate elliptic systems with right hand side in a Marcinkiewicz space[J]. Mathematics in Engineering, 2023, 5(3): 1-23. doi: 10.3934/mine.2023055







 DownLoad:
DownLoad: