The aim of the current study was to explore the gene enrichment and dysregulated pathways on the basis of interaction network analysis of SLC30A8 in type 1 diabetes mellitus (T1DM). SLC30A8 polymorphism could be characterized as a beneficial tool to identify the interacting gene in developing T1DM.
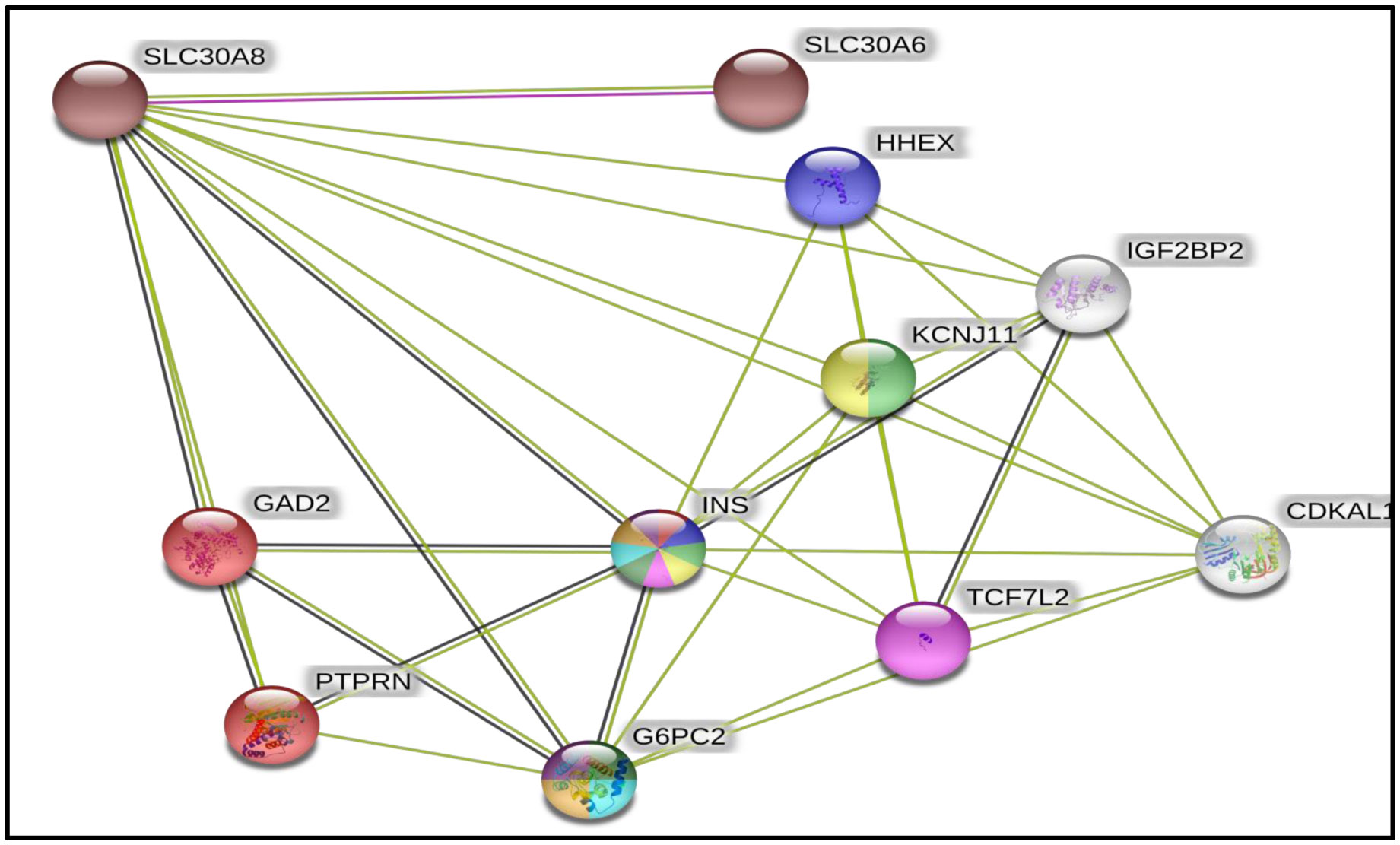
SLC30A8 interacting protein interaction network was obtained by String Interaction network Version 11.0. Ten proteins were identified interacting with SLC30A8 and were analysed by protein-protein interaction and enrichment network analysis along with Functional Enrichment analysis tool (FunRich 3.1.3) to map the gene data sets. In entire analysis, FunRich database was used as background against all annotated gene/protein list. Protein-protein interaction (PPI) and enrichment network analysis of the selected protein: SLC30A8 gene along with gene mapping and pathway enrichment were performed using FunRich 3.1.3 and String Interaction network Version 11.0.
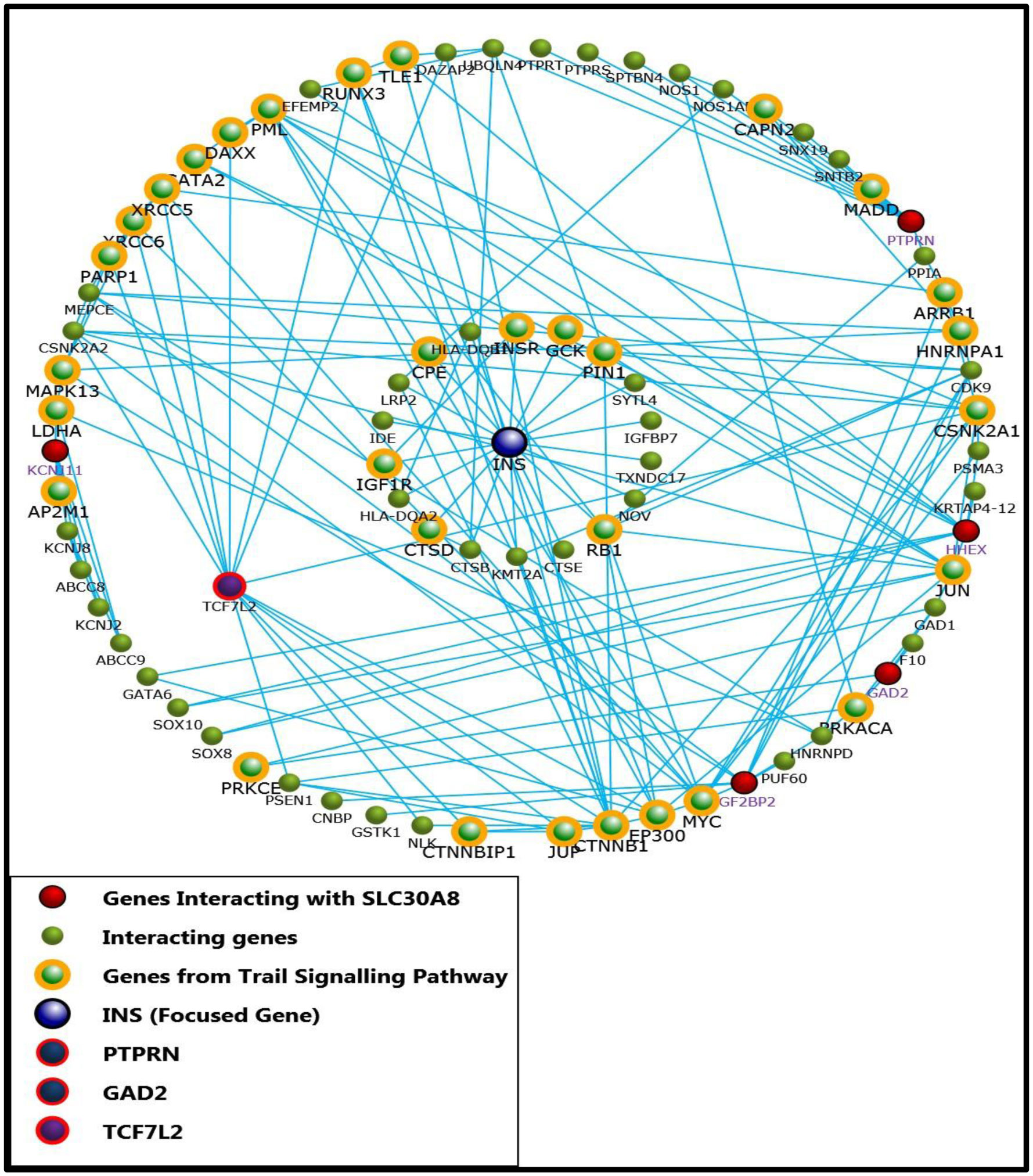
Biological pathway grouping displayed enriched proteins in TRAIL signalling pathway (p < 0.001). PTPRN, GAD2 and TCF7L2 were enriched in TRAIL Signalling pathway when INS was made focused gene and directly interacting with SLC30A8.
TRAIL signalling pathways were enriched in T1DM. Therefore, SLC30A8 along with PTPRN, GAD2 and TCF7L2 involved in TRAIL pathway must be further explored to understand their in vivo role in T1DM.
Citation: Afreen Bhatty, Zile Rubab, Hafiz Syed Mohammad Osama Jafri, Sheh Zano. Identification of dysregulated pathways through SLC30A8 protein interaction in type 1 diabetes mellitus[J]. AIMS Molecular Science, 2021, 8(4): 301-310. doi: 10.3934/molsci.2021023
The aim of the current study was to explore the gene enrichment and dysregulated pathways on the basis of interaction network analysis of SLC30A8 in type 1 diabetes mellitus (T1DM). SLC30A8 polymorphism could be characterized as a beneficial tool to identify the interacting gene in developing T1DM.
SLC30A8 interacting protein interaction network was obtained by String Interaction network Version 11.0. Ten proteins were identified interacting with SLC30A8 and were analysed by protein-protein interaction and enrichment network analysis along with Functional Enrichment analysis tool (FunRich 3.1.3) to map the gene data sets. In entire analysis, FunRich database was used as background against all annotated gene/protein list. Protein-protein interaction (PPI) and enrichment network analysis of the selected protein: SLC30A8 gene along with gene mapping and pathway enrichment were performed using FunRich 3.1.3 and String Interaction network Version 11.0.
Biological pathway grouping displayed enriched proteins in TRAIL signalling pathway (p < 0.001). PTPRN, GAD2 and TCF7L2 were enriched in TRAIL Signalling pathway when INS was made focused gene and directly interacting with SLC30A8.
TRAIL signalling pathways were enriched in T1DM. Therefore, SLC30A8 along with PTPRN, GAD2 and TCF7L2 involved in TRAIL pathway must be further explored to understand their in vivo role in T1DM.

| [1] |
Roep BO, Thomaidou S, van Tienhoven R, et al. (2020) Type 1 diabetes mellitus as a disease of the β-cell (do not blame the immune system?). Nat Rev Endocrinol 17: 150-161. doi: 10.1038/s41574-020-00443-4

|
| [2] |
Bhatty A, Baig S, Shahid MA (2019) Emerging role of zinc transporter-8 autoantibodies (ZnT8A) in type 1 diabetes mellitus-a review. J Adv Med Med Res 1-10. doi: 10.9734/jammr/2019/v31i630304
|
| [3] |
Shera AS, Miyan Z, Basit A, et al. (2008) Trends of type 1 diabetes in Karachi, Pakistan. Pediatr Diabetes 9: 401-406. doi: 10.1111/j.1399-5448.2008.00309.x
|
| [4] |
Gomes KF, Semzezem C, Batista R, et al. (2017) Importance of zinc transporter 8 autoantibody in the diagnosis of type 1 diabetes in Latin Americans. Sci Rep 7: 1-7. doi: 10.1038/s41598-016-0028-x
|
| [5] |
Regnell SE, Lernmark Å (2017) Early prediction of autoimmune (type 1) diabetes. Diabetologia 60: 1370-1381. doi: 10.1007/s00125-017-4308-1
|
| [6] |
Wan H, Merriman C, Atkinson MA, et al. (2017) Proteoliposome-based full-length ZnT8 self-antigen for type 1 diabetes diagnosis on a plasmonic platform. Proc Natl Acad Sci U S A 114: 10196-10201. doi: 10.1073/pnas.1711169114
|
| [7] |
Thirunavukkarasu R, Asirvatham AJ, Chitra A (2018) SLC30A8 Gene rs13266634 C/T Polymorphism in Type 1 Diabetes Children of Tamil Nadu, India. J Clin Res Pediatr Endocrinol 11: 55-60. doi: 10.4274/jcrpe.galenos.2018.2018.0195
|
| [8] |
Braga-Neto UM, Dougherty ER (2004) Is cross-validation valid for small-sample microarray classification? Bioinformatics 20: 374-380. doi: 10.1093/bioinformatics/btg419
|
| [9] | Shi Z, Yu X, Wang X, et al. (2016) Identification of dysregulated pathways involved in hepatitis C based on network pathway enrichment analysis. Int J Clin Exp Pathol 9: 5611-5619. |
| [10] |
Abatangelo L, Maglietta R, Distaso A, et al. (2009) Comparative study of gene set enrichment methods. BMC Bioinf 10: 275. doi: 10.1186/1471-2105-10-275
|
| [11] |
Huang DW, Sherman BT, Lempicki RA (2009) Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res 37: 1-3. doi: 10.1093/nar/gkn923
|
| [12] |
Liu Y, Koyutürk M, Barnholtz-Sloan JS, et al. (2012) Gene interaction enrichment and network analysis to identify dysregulated pathways and their interactions in complex diseases. BMC Syst Biol 6: 65. doi: 10.1186/1752-0509-6-65
|
| [13] |
Glaab E, Baudot A, Krasnogor N, et al. (2012) EnrichNet: network-based gene set enrichment analysis. Bioinformatics 28: i451-i457. doi: 10.1093/bioinformatics/bts389
|
| [14] | Bhatty A, Baig S, Fawwad A, et al. (2020) Association of zinc transporter-8 autoantibody (ZnT8A) with Type 1 diabetes mellitus. Cureus 12: e7263. |
| [15] | STRING, Protein-protein interaction networks functional enrichment analysis Available from: https://string-db.org/. |
| [16] | FunRich: Functional Enrichment analysis tool Available from: http://www.funrich.org/. |
| [17] |
Kahaly GJ, Hansen MP (2016) Type 1 diabetes associated autoimmunity. Autoimmun Rev 15: 644-648. doi: 10.1016/j.autrev.2016.02.017
|
| [18] |
Maares M, Haase H (2016) Zinc and immunity: An essential interrelation. Arch Biochem Biophys 611: 58-65. doi: 10.1016/j.abb.2016.03.022
|
| [19] |
Ruan X, Li H, Liu BO, et al. (2015) A novel method to identify pathways associated with renal cell carcinoma based on a gene co-expression network. Oncol Rep 34: 567-576. doi: 10.3892/or.2015.4038
|
| [20] |
Sanlioglu AD, Griffith TS, Omer A, et al. (2008) Molecular mechanisms of death ligand-mediated immune modulation: A gene therapy model to prolong islet survival in type 1 diabetes. J Cell Biochem 104: 710-720. doi: 10.1002/jcb.21677
|
| [21] |
Kang S, Park EJ, Joe Y, et al. (2010) Systemic delivery of TNF-related apoptosis-inducing ligand (TRAIL) elevates levels of tissue inhibitor of metalloproteinase-1 (TIMP-1) and prevents type 1 diabetes in nonobese diabetic mice. Endocrinology 151: 5638-5646. doi: 10.1210/en.2009-0478
|
| [22] |
Riquelme Medina I, Lubovac-Pilav Z (2016) Gene co-expression network analysis for identifying modules and functionally enriched pathways in type 1 diabetes. Plos One 11: e0156006. doi: 10.1371/journal.pone.0156006
|
| [23] |
De Jong VM, Van der Slik AR, Laban S, et al. (2016) Survival of autoreactive T lymphocytes by microRNA-mediated regulation of apoptosis through TRAIL and Fas in type 1 diabetes. Genes Immun 17: 342-348. doi: 10.1038/gene.2016.29
|
| [24] |
Xie J, Zhang B, Lan MS, et al. (1998) Genomic structure and promoter sequence of the insulin-dependent diabetes mellitus autoantigen, IA-2 (PTPRN). Genomics 54: 338-343. doi: 10.1006/geno.1998.5583
|
| [25] |
Redondo MJ, Geyer S, Steck AK, et al. (2018) Type 1 diabetes TrialNet study group. TCF7L2 genetic variants contribute to phenotypic heterogeneity of type 1 diabetes. Diabetes Care 41: 311-317. doi: 10.2337/dc17-0961
|
| [26] |
Sanlioglu AD, Dirice E, Elpek O, et al. (2008) High levels of endogenous tumor necrosis factor-related apoptosis-inducing ligand expression correlate with increased cell death in human pancreas. Pancreas 36: 385-393. doi: 10.1097/MPA.0b013e318158a4e5
|
Figures(2) / Tables(1)

Afreen Bhatty, Zile Rubab, Hafiz Syed Mohammad Osama Jafri, Sheh Zano. Identification of dysregulated pathways through SLC30A8 protein interaction in type 1 diabetes mellitus[J]. AIMS Molecular Science, 2021, 8(4): 301-310. doi: 10.3934/molsci.2021023







 DownLoad:
DownLoad: