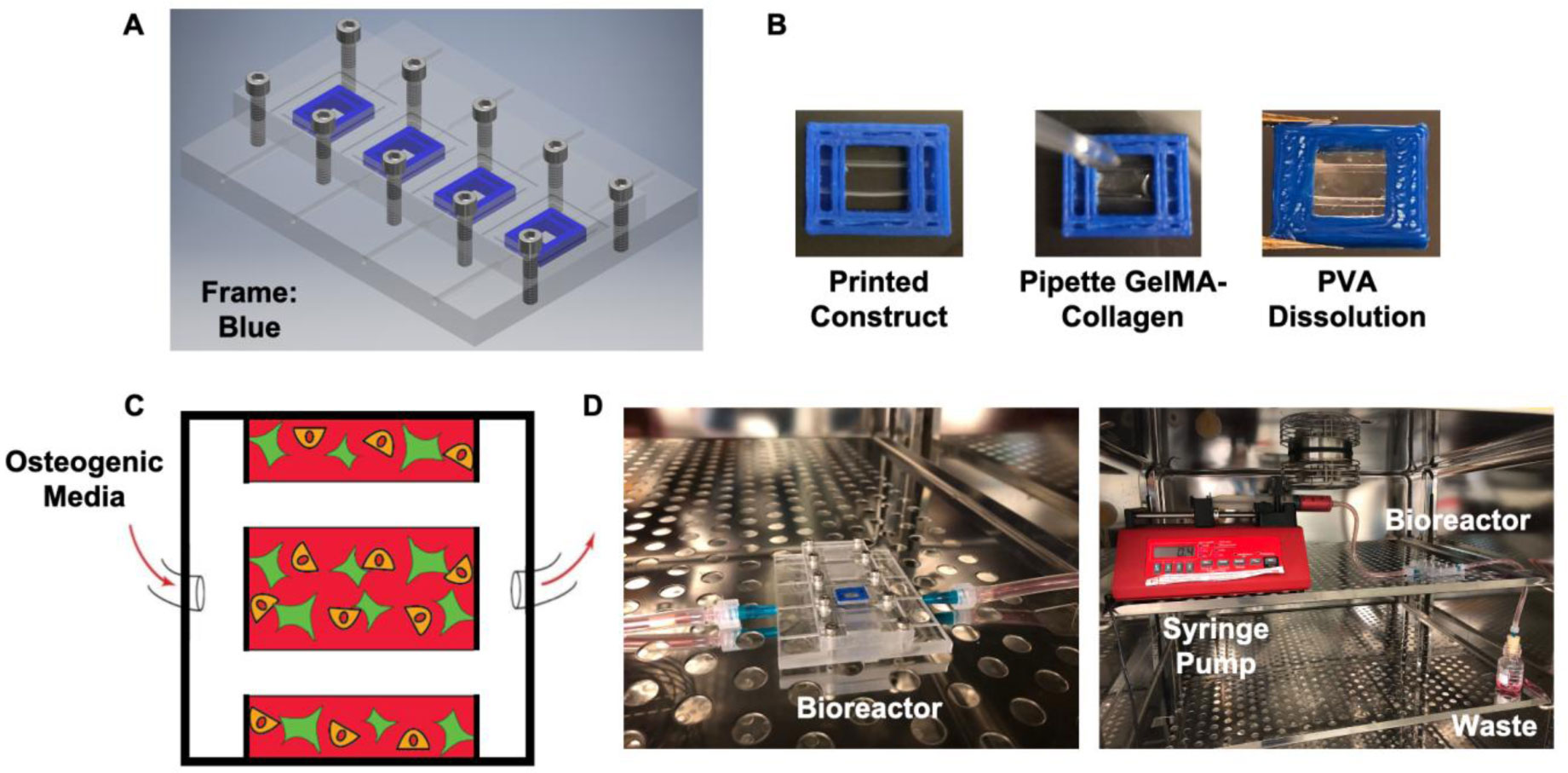
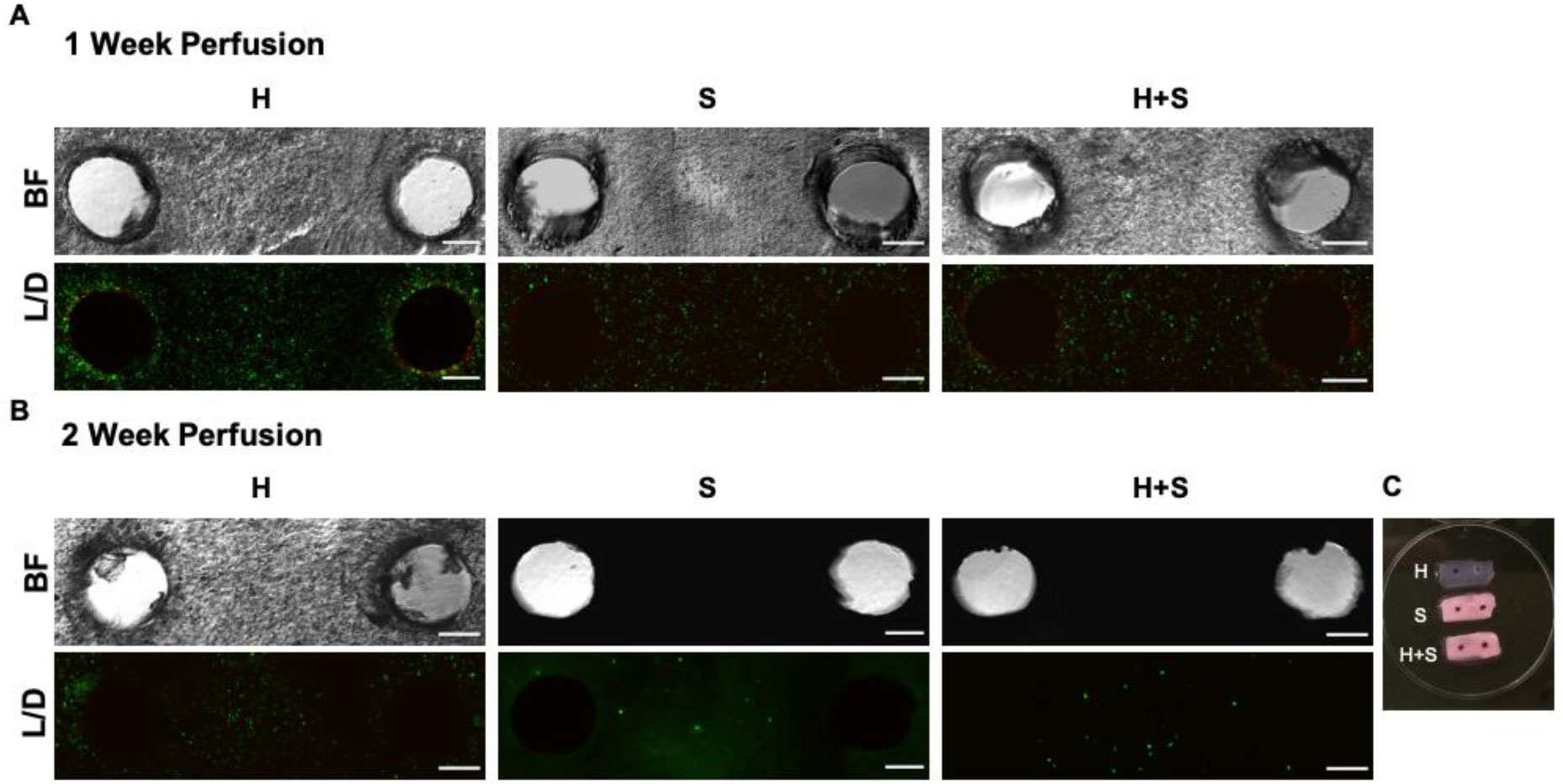
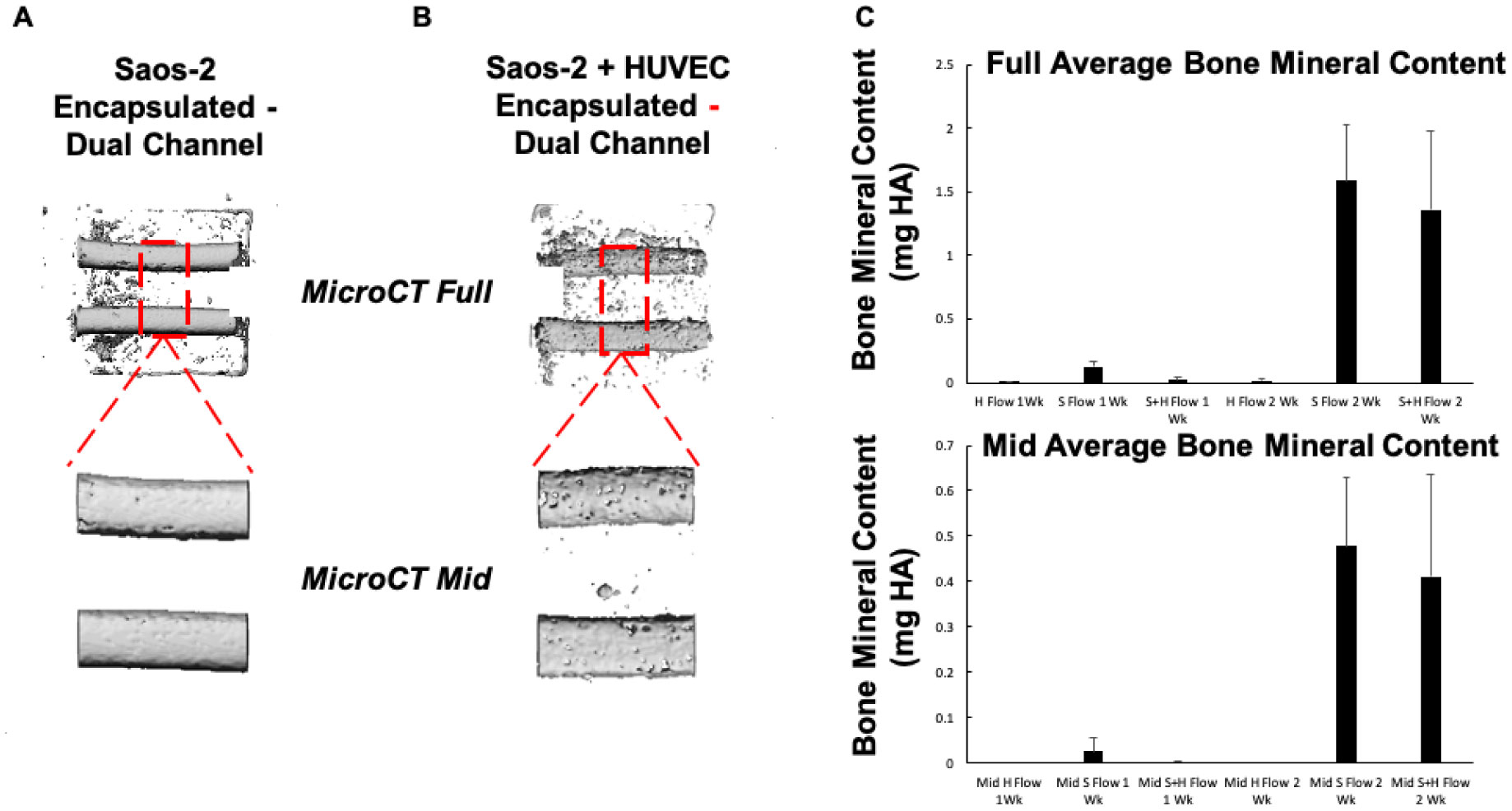
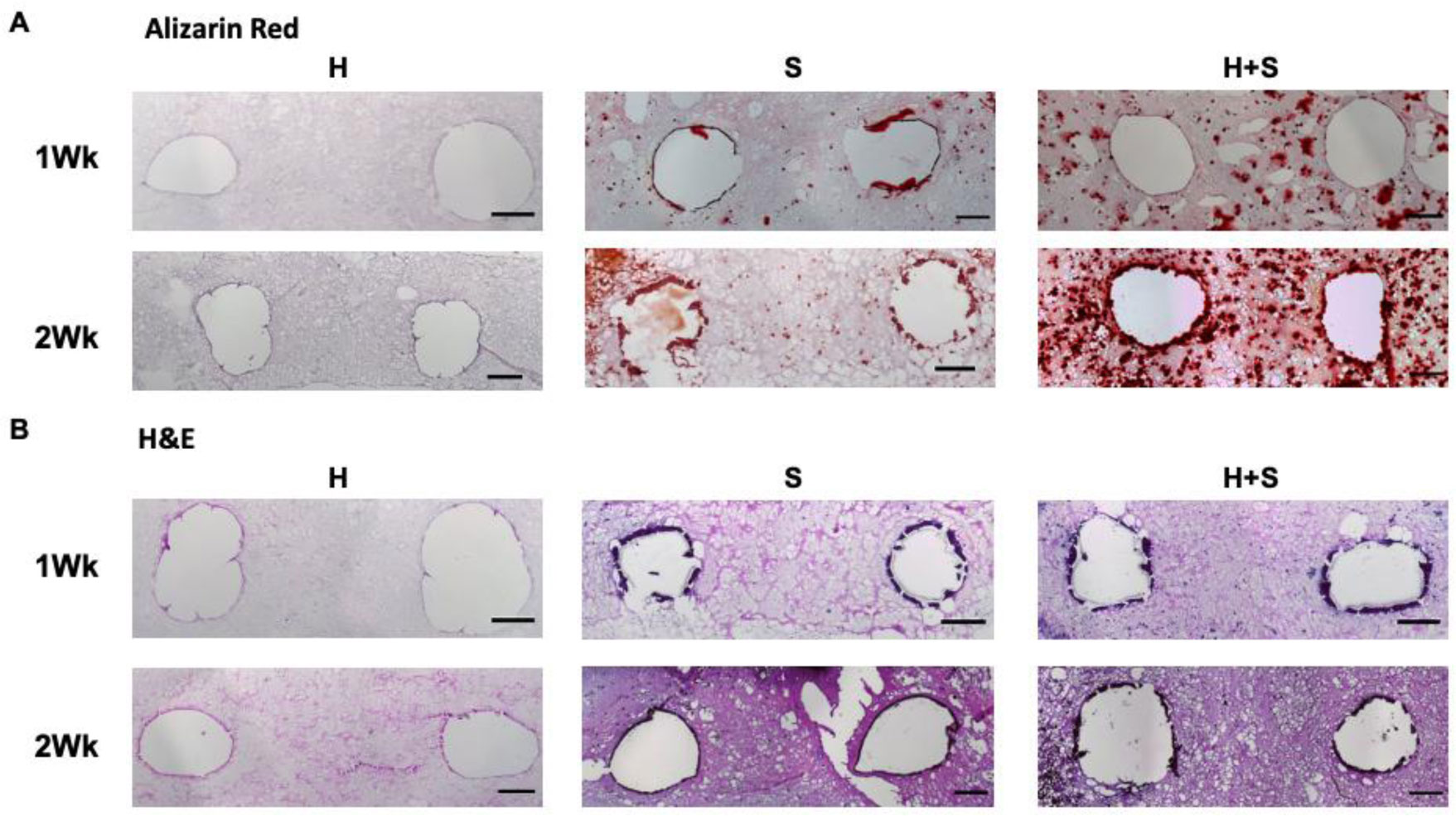
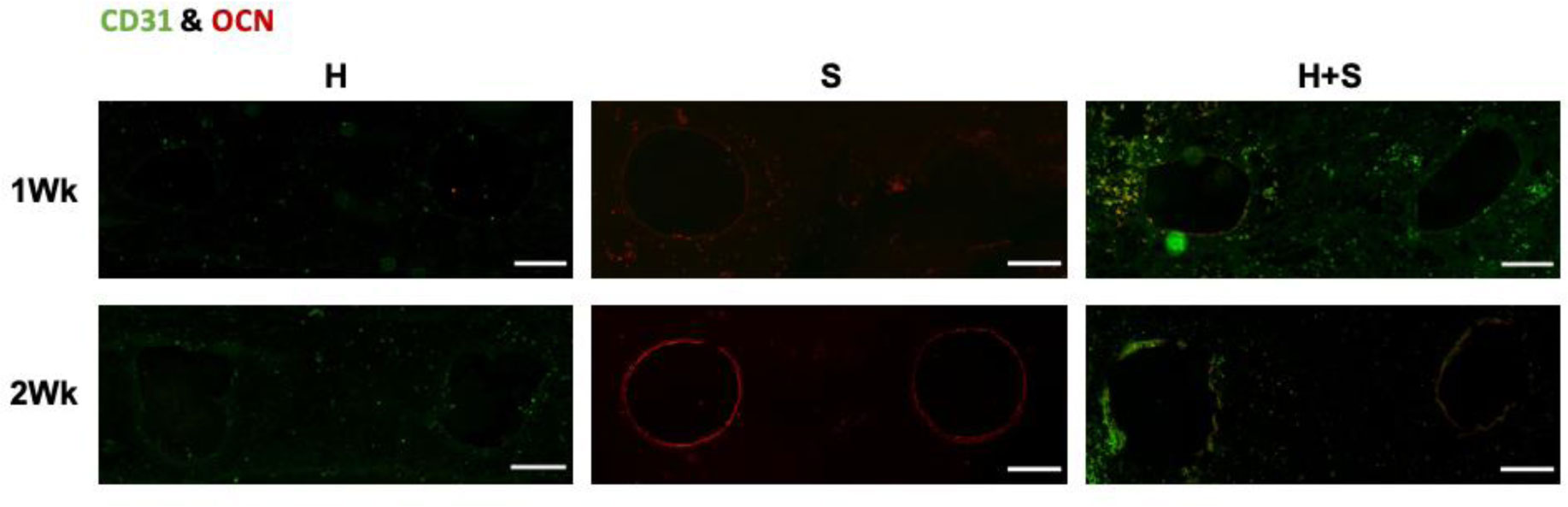
In this work, we report on a perfusion-based co-culture system that could be used for bone tissue engineering applications. The model system is created using a combination of Primary Human Umbilical Vein Endothelial Cells (HUVECs) and osteoblast-like Saos-2 cells encapsulated within a Gelatin Methacrylate (GelMA)-collagen hydrogel blend contained within 3D printed, perfusable constructs. The constructs contain dual channels, within a custom-built bioreactor, that were perfused with osteogenic media for up to two weeks in order to induce mineral deposition. Mineral deposition in constructs containing only HUVECs, only Saos-2 cells, or a combination thereof was quantified by microCT to determine if the combination of endothelial cells and bone-like cells increased mineral deposition. Histological and fluorescent staining was used to verify mineral deposition and cellular function both along and between the perfused channels. While there was not a quantifiable difference in the amount of mineral deposited in Saos-2 only versus Saos-2 plus HUVEC samples, the location of the deposited mineral differed dramatically between the groups and indicated that the addition of HUVECs within the GelMA matrix allowed Saos-2 cells, in diffusion limited regions of the construct, to deposit bone mineral. This work serves as a model on how to create perfusable bone tissue engineering constructs using a combination of 3D printing and cellular co-cultures.
Citation: Stephen W. Sawyer, Kairui Zhang, Jason A. Horton, Pranav Soman. Perfusion-based co-culture model system for bone tissue engineering[J]. AIMS Bioengineering, 2020, 7(2): 91-105. doi: 10.3934/bioeng.2020009
In this work, we report on a perfusion-based co-culture system that could be used for bone tissue engineering applications. The model system is created using a combination of Primary Human Umbilical Vein Endothelial Cells (HUVECs) and osteoblast-like Saos-2 cells encapsulated within a Gelatin Methacrylate (GelMA)-collagen hydrogel blend contained within 3D printed, perfusable constructs. The constructs contain dual channels, within a custom-built bioreactor, that were perfused with osteogenic media for up to two weeks in order to induce mineral deposition. Mineral deposition in constructs containing only HUVECs, only Saos-2 cells, or a combination thereof was quantified by microCT to determine if the combination of endothelial cells and bone-like cells increased mineral deposition. Histological and fluorescent staining was used to verify mineral deposition and cellular function both along and between the perfused channels. While there was not a quantifiable difference in the amount of mineral deposited in Saos-2 only versus Saos-2 plus HUVEC samples, the location of the deposited mineral differed dramatically between the groups and indicated that the addition of HUVECs within the GelMA matrix allowed Saos-2 cells, in diffusion limited regions of the construct, to deposit bone mineral. This work serves as a model on how to create perfusable bone tissue engineering constructs using a combination of 3D printing and cellular co-cultures.

| [1] |
Buckwalter JA, Glimcher MJ, Cooper RR (1995) Bone biology. Part I: Structure, blood supply, cells, matrix, and mineralization. J Bone Joint Surg 77: 1256-1275. doi: 10.2106/00004623-199508000-00019

|
| [2] | McCarthy I (2006) The physiology of bone blood flow: a review. JB & JS 88: 4-9. |
| [3] |
Laroche M (2002) Intraosseous circulation from physiology to disease. Joint Bone Spine 69: 262-269. doi: 10.1016/S1297-319X(02)00391-3
|
| [4] |
Mercado-Pagán ÁE, Stahl AM, Shanjani Y, et al. (2015) Vascularization in bone tissue engineering constructs. Ann Biomed Eng 43: 718-729. doi: 10.1007/s10439-015-1253-3
|
| [5] |
Rouwkema J, Westerweel PE, De Boer J, et al. (2009) The use of endothelial progenitor cells for prevascularized bone tissue engineering. Tissue Eng Part A 15: 2015-2027. doi: 10.1089/ten.tea.2008.0318
|
| [6] |
Krishnan L, Willett NJ, Guldberg RE (2014) Vascularization strategies for bone regeneration. Ann Biomed Eng 42: 432-444. doi: 10.1007/s10439-014-0969-9
|
| [7] |
Shanjani Y, Kang Y, Zarnescu L, et al. (2017) Endothelial pattern formation in hybrid constructs of additive manufactured porous rigid scaffolds and cell-laden hydrogels for orthopedic applications. J Mech Behav Biomed Mater 65: 356-372. doi: 10.1016/j.jmbbm.2016.08.037
|
| [8] |
Murphy WL, Peters MC, Kohn DH, et al. (2000) Sustained release of vascular endothelial growth factor from mineralized poly (lactide-co-glycolide) scaffolds for tissue engineering. Biomaterials 21: 2521-2527. doi: 10.1016/S0142-9612(00)00120-4
|
| [9] |
Lee KY, Peters MC, Anderson KW, et al. (2000) Controlled growth factor release from synthetic extracellular matrices. Nature 408: 998-1000. doi: 10.1038/35050141
|
| [10] |
Sheridan MH, Shea LD, Peters MC, et al. (2000) Bioabsorbable polymer scaffolds for tissue engineering capable of sustained growth factor delivery. J Controll Release 64: 91-102. doi: 10.1016/S0168-3659(99)00138-8
|
| [11] |
Peters MC, Polverini PJ, Mooney DJ (2002) Engineering vascular networks in porous polymer matrices. J Biomed Mater Res A 60: 668-678. doi: 10.1002/jbm.10134
|
| [12] |
Wang L, Fan H, Zhang ZY, et al. (2010) Osteogenesis and angiogenesis of tissue-engineered bone constructed by prevascularized β-tricalcium phosphate scaffold and mesenchymal stem cells. Biomaterials 31: 9452-9461. doi: 10.1016/j.biomaterials.2010.08.036
|
| [13] |
Yu H, VandeVord PJ, Gong W, et al. (2008) Promotion of osteogenesis in tissue-engineered bone by pre-seeding endothelial progenitor cells-derived endothelial cells. J Orthop Res 26: 1147-1152. doi: 10.1002/jor.20609
|
| [14] |
Villars F, Bordenave L, Bareille R, et al. (2000) Effect of human endothelial cells on human bone marrow stromal cell phenotype: role of VEGF? J Cell Biochem 79: 672-685. doi: 10.1002/1097-4644(20001215)79:4<672::AID-JCB150>3.0.CO;2-2
|
| [15] |
Santos MI, Reis RL (2010) Vascularization in bone tissue engineering: physiology, current strategies, major hurdles and future challenges. Macromol Biosci 10: 12-27. doi: 10.1002/mabi.200900107
|
| [16] |
Villars F, Guillotin B, Amedee T, et al. (2002) Effect of HUVEC on human osteoprogenitor cell differentiation needs heterotypic gap junction communication. Am J Physiol-Cell Ph 282: C775-C85. doi: 10.1152/ajpcell.00310.2001
|
| [17] |
Stegen S, van Gastel N, Carmeliet G (2015) Bringing new life to damaged bone: the importance of angiogenesis in bone repair and regeneration. Bone 70: 19-27. doi: 10.1016/j.bone.2014.09.017
|
| [18] |
Ghanaati S, Fuchs S, Webber MJ, et al. (2011) Rapid vascularization of starch–poly (caprolactone) in vivo by outgrowth endothelial cells in co-culture with primary osteoblasts. J Tissue Eng Regen M 5: e136-e143. doi: 10.1002/term.373
|
| [19] |
Guillotin B, Bareille R, Bourget C, et al. (2008) Interaction between human umbilical vein endothelial cells and human osteoprogenitors triggers pleiotropic effect that may support osteoblastic function. Bone 42: 1080-1091. doi: 10.1016/j.bone.2008.01.025
|
| [20] |
Unger RE, Sartoris A, Peters K, et al. (2007) Tissue-like self-assembly in cocultures of endothelial cells and osteoblasts and the formation of microcapillary-like structures on three-dimensional porous biomaterials. Biomaterials 28: 3965-3976. doi: 10.1016/j.biomaterials.2007.05.032
|
| [21] | Ma JL, van den Beucken JJJP, Yang F, et al. (2011) Coculture of osteoblasts and endothelial cells: optimization of culture medium and cell ratio. Tissue Eng Part C 17: 349-357. |
| [22] |
Black AF, Berthod F, L'Heureux N, et al. (1998) In vitro reconstruction of a human capillary-like network in a tissue-engineered skin equivalent. FASEB J 12: 1331-1340. doi: 10.1096/fasebj.12.13.1331
|
| [23] |
Chiu LLY, Montgomery M, Liang Y, et al. (2012) Perfusable branching microvessel bed for vascularization of engineered tissues. Proc Natl Acad Sci USA 109: E3414-E3423. doi: 10.1073/pnas.1210580109
|
| [24] |
Zheng Y, Chen J, Craven M, et al. (2012) In vitro microvessels for the study of angiogenesis and thrombosis. Proc Natl Acad Sci USA 109: 9342-9347. doi: 10.1073/pnas.1201240109
|
| [25] |
Chen YC, Lin RZ, Qi H, et al. (2012) Functional human vascular network generated in photocrosslinkable gelatin methacrylate hydrogels. Adv Funct Mater 22: 2027-2039. doi: 10.1002/adfm.201101662
|
| [26] |
Cuchiara MP, Gould DJ, McHale MK, et al. (2012) Integration of self-assembled microvascular networks with microfabricated PEG-based hydrogels. Adv Funct Mater 22: 4511-4518. doi: 10.1002/adfm.201200976
|
| [27] |
Chrobak KM, Potter DR, Tien J (2006) Formation of perfused, functional microvascular tubes in vitro. Microvasc Res 71: 185-196. doi: 10.1016/j.mvr.2006.02.005
|
| [28] |
Price GM, Wong KHK, Truslow JG, et al. (2010) Effect of mechanical factors on the function of engineered human blood microvessels in microfluidic collagen gels. Biomaterials 31: 6182-6189. doi: 10.1016/j.biomaterials.2010.04.041
|
| [29] |
Nichol JW, Koshy ST, Bae H, et al. (2010) Cell-laden microengineered gelatin methacrylate hydrogels. Biomaterials 31: 5536-5544. doi: 10.1016/j.biomaterials.2010.03.064
|
| [30] | Park JH, Chung BG, Lee WG, et al. (2010) Microporous cell-laden hydrogels for engineered tissue constructs. Biotechnol Bioeng 106: 138-148. |
| [31] |
Sadr N, Zhu M, Osaki T, et al. (2011) SAM-based cell transfer to photopatterned hydrogels for microengineering vascular-like structures. Biomaterials 32: 7479-7490. doi: 10.1016/j.biomaterials.2011.06.034
|
| [32] |
Therriault D, White SR, Lewis JA (2003) Chaotic mixing in three-dimensional microvascular networks fabricated by direct-write assembly. Nat Mater 2: 265-271. doi: 10.1038/nmat863
|
| [33] |
Golden AP, Tien J (2007) Fabrication of microfluidic hydrogels using molded gelatin as a sacrificial element. Lab Chip 7: 720-725. doi: 10.1039/b618409j
|
| [34] |
Miller JS, Stevens KR, Yang MT, et al. (2012) Rapid casting of patterned vascular networks for perfusable engineered three-dimensional tissues. Nat Mater 11: 768-774. doi: 10.1038/nmat3357
|
| [35] |
Annabi N, Tamayol A, Uquillas JA, et al. (2014) 25th anniversary article: Rational design and applications of hydrogels in regenerative medicine. Adv Mater 26: 85-124. doi: 10.1002/adma.201303233
|
| [36] |
Bertassoni LE, Cardoso JC, Manoharan V, et al. (2014) Direct-write bioprinting of cell-laden methacrylated gelatin hydrogels. Biofabrication 6: 024105. doi: 10.1088/1758-5082/6/2/024105
|
| [37] |
Tan Y, Richards DJ, Trusk TC, et al. (2014) 3D printing facilitated scaffold-free tissue unit fabrication. Biofabrication 6: 024111. doi: 10.1088/1758-5082/6/2/024111
|
| [38] |
Wu W, DeConinck A, Lewis JA (2011) Omnidirectional printing of 3D microvascular networks. Adv Mater 23: H178-H183. doi: 10.1002/adma.201004625
|
| [39] |
Lee VK, Kim DY, Ngo H, et al. (2014) Creating perfused functional vascular channels using 3D bio-printing technology. Biomaterials 35: 8092-8102. doi: 10.1016/j.biomaterials.2014.05.083
|
| [40] |
Miller JS, Stevens KR, Yang MT, et al. (2012) Rapid casting of patterned vascular networks for perfusable engineered three-dimensional tissues. Nat Mater 11: 768-774. doi: 10.1038/nmat3357
|
| [41] |
Kolesky DB, Truby RL, Gladman AS, et al. (2014) 3D bioprinting of vascularized, heterogeneous cell-laden tissue constructs. Adv Mater 26: 3124-3130. doi: 10.1002/adma.201305506
|
| [42] |
Skardal A, Zhang J, McCoard L, et al. (2010) Photocrosslinkable hyaluronan-gelatin hydrogels for two-step bioprinting. Tissue Eng Part A 16: 2675-2685. doi: 10.1089/ten.tea.2009.0798
|
| [43] |
Gao Q, He Y, Fu J-z, et al. (2015) Coaxial nozzle-assisted 3D bioprinting with built-in microchannels for nutrients delivery. Biomaterials 61: 203-215. doi: 10.1016/j.biomaterials.2015.05.031
|
| [44] |
Yang L, Shridhar SV, Gerwitz M, et al. (2016) An in vitro vascular chip using 3D printing-enabled hydrogel casting. Biofabrication 8: 035015. doi: 10.1088/1758-5090/8/3/035015
|
| [45] |
Tocchio A, Tamplenizza M, Martello F, et al. (2015) Versatile fabrication of vascularizable scaffolds for large tissue engineering in bioreactor. Biomaterials 45: 124-131. doi: 10.1016/j.biomaterials.2014.12.031
|
| [46] |
Sawyer SW, Shridhar SV, Zhang K, et al. (2018) Perfusion directed 3D mineral formation within cell-laden hydrogels. Biofabrication 10: 035013. doi: 10.1088/1758-5090/aacb42
|
| [47] |
Sladkova M, De Peppo GM (2014) Bioreactor systems for human bone tissue engineering. Processes 2: 494-525. doi: 10.3390/pr2020494
|
| [48] |
Rice JJ, Martino MM, De Laporte L, et al. (2013) Engineering the regenerative microenvironment with biomaterials. Adv Healthc Mater 2: 57-71. doi: 10.1002/adhm.201200197
|
| [49] |
Cartmell SH, Porter BD, García AJ, et al. (2003) Effects of medium perfusion rate on cell-seeded three-dimensional bone constructs in vitro. Tissue Eng 9: 1197-1203. doi: 10.1089/10763270360728107
|
| [50] |
Albrecht LD, Sawyer SW, Soman P (2016) Developing 3D scaffolds in the field of tissue engineering to treat complex bone defects. 3D Print Addit Manuf 3: 106-112. doi: 10.1089/3dp.2016.0006
|
| [51] |
Hutmacher DW (2001) Scaffold design and fabrication technologies for engineering tissues—state of the art and future perspectives. J Biomater Sci Poly Ed 12: 107-124. doi: 10.1163/156856201744489
|
| [52] |
Burg KJL, Porter S, Kellam JF (2000) Biomaterial developments for bone tissue engineering. Biomaterials 21: 2347-2359. doi: 10.1016/S0142-9612(00)00102-2
|
| [53] |
Reichert JC, Hutmacher DW (2011) Bone tissue engineering. Tissue Engineering Heidelberg: Springer, 431-456. doi: 10.1007/978-3-642-02824-3_21
|
| [54] |
Bose S, Vahabzadeh S, Bandyopadhyay A (2013) Bone tissue engineering using 3D printing. Mater Today 16: 496-504. doi: 10.1016/j.mattod.2013.11.017
|
| [55] |
Stevens MM (2008) Biomaterials for bone tissue engineering. Mater Today 11: 18-25. doi: 10.1016/S1369-7021(08)70086-5
|
| [56] |
Baranski JD, Chaturvedi RR, Stevens KR, et al. (2013) Geometric control of vascular networks to enhance engineered tissue integration and function. Proc Natl Acad Sci USA 110: 7586-7591. doi: 10.1073/pnas.1217796110
|
| [57] |
Lee VK, Lanzi AM, Ngo H, et al. (2014) Generation of multi-scale vascular network system within 3D hydrogel using 3D bio-printing technology. Cell Mol Bioeng 7: 460-472. doi: 10.1007/s12195-014-0340-0
|
| [58] |
Sawyer S, Oest M, Margulies B, et al. (2016) Behavior of encapsulated saos-2 cells within gelatin methacrylate hydrogels. J Tissue Sci Eng 7: 1000173. doi: 10.4172/2157-7552.1000173
|
| [59] |
Sawyer SW, Dong P, Venn S, et al. (2017) Conductive gelatin methacrylate-poly (aniline) hydrogel for cell encapsulation. Biomed Phys Eng Express 4: 015005. doi: 10.1088/2057-1976/aa91f9
|
| [60] |
Mikos AG, Sarakinos G, Lyman MD, et al. (1993) Prevascularization of porous biodegradable polymers. Biotechnol Bioeng 42: 716-723. doi: 10.1002/bit.260420606
|
 bioeng-07-02-009-s001.pdf bioeng-07-02-009-s001.pdf |
 |
Figures(5)

Stephen W. Sawyer, Kairui Zhang, Jason A. Horton, Pranav Soman. Perfusion-based co-culture model system for bone tissue engineering[J]. AIMS Bioengineering, 2020, 7(2): 91-105. doi: 10.3934/bioeng.2020009







 DownLoad:
DownLoad: