Citation: Akira Nishimura, Tadaki Inoue, Yoshito Sakakibara, Masafumi Hirota, Akira Koshio, Fumio Kokai, Eric Hu. Optimum molar ratio of H2 and H2O to reduce CO2 using Pd/TiO2[J]. AIMS Materials Science, 2019, 6(4): 464-483. doi: 10.3934/matersci.2019.4.464

| [1] | Greenhouse Gases Observing Satellite GOSAT "IBUSUKI", 2019. Available from: http://www.gosat.nies.go.jp/en/. |
| [2] |
Tahir M, Amin NS (2015) Indium-doped TiO2 nanoparticles for photocatalytic CO2 reduction with H2O vapors to CH4. Appl Catal B-Environ 162: 98–109. doi: 10.1016/j.apcatb.2014.06.037

|
| [3] |
Abdulah H, Khan MMR, Ong HR, et al. (2017) Modified TiO2 photocatalyst for CO2 photocatalytic reduction: an overview. J CO2 Util 22: 15–32. doi: 10.1016/j.jcou.2017.08.004
|
| [4] |
Sohn Y, Huang W, Taghipour F (2017) Recent progress and perspectives in the photocatalytic CO2 reduction of Ti-oxide-based nanomaterials. Appl Surf Sci 396: 1696–1711. doi: 10.1016/j.apsusc.2016.11.240
|
| [5] |
Nahar S, Zain MF, Kadhum AAH, et al. (2017) Advances in photocatalytic CO2 reduction with water: A Review. Materials 10: 629. doi: 10.3390/ma10060629
|
| [6] |
Ola O, Maroto-Valer MM (2015) Review of material design and reactor engineering on TiO2 photocatalysis for CO2 reduction. J Photoch Photobio C 24: 16–42. doi: 10.1016/j.jphotochemrev.2015.06.001
|
| [7] |
Xie S, Wang Y, Zhang Q, et al. (2014) MgO- and Pt-promoted TiO2 as an efficient photocatalyst for the prefential reduction of carbon dioxide in the presence of water. ACS Catal 4: 3644–3653. doi: 10.1021/cs500648p
|
| [8] |
Khalid NR, Ahmed E, Niaz NA, et al. (2017) Highly visible light responsive metal loaded N/TiO2 nanoparticles for photocatalytic conversion of CO2 into methane. Ceram Int 43: 6771–6777. doi: 10.1016/j.ceramint.2017.02.093
|
| [9] | Tan LL, Ong WJ, Chai SP, et al. (2015) Noble metal modified reduced graphene oxide/TiO2 ternary nanostructures for efficient visible-light-driven photoreduction of carbon dioxide into methane. Appl Catal B-Environ 166–167: 251–259. |
| [10] |
Cmarillo R, Toston S, Martinez F, et al. (2018) Improving the photo-reduction of CO2 to fuels with catalysts synthesized under high pressure: Cu/TiO2. J Chem Technol Biot 93: 1237–1248. doi: 10.1002/jctb.5477
|
| [11] |
Aguirre ME, Zhou R, Eugene AJ, et al. (2017) Cu2O/TiO2 heterostructures for CO2 reduction through a direct z-scheme: protecting Cu2O from photocorrosion. Appl Catal B-Environ 217: 485–493. doi: 10.1016/j.apcatb.2017.05.058
|
| [12] |
Marci G, Garcia-Lopez EI, Palmisano L (2014) Photocatalytic CO2 reduction in gas-solid regime in the presence of H2O by using GaP/TiO2 composite as photocatalyst under simulated solar light. Catal Commun 53: 38–41. doi: 10.1016/j.catcom.2014.04.024
|
| [13] |
Beigi AA, Fatemi S, Salehi Z (2014) Synthesis of nanocomposite CdS/TiO2 and investigation of its photocatalytic activity of CO2 reduction to CO and CH4 under visible light irradiation. J CO2 Util 7: 23–29. doi: 10.1016/j.jcou.2014.06.003
|
| [14] |
Fang Z, Li S, Gong Y, et al. (2014) Comparison of catalytic activity of carbon-based AgBr nanocomposites for conversion of CO2 under visible light. J Saudi Chem Soc 18: 299–307. doi: 10.1016/j.jscs.2013.08.003
|
| [15] |
Xu F, Zhang J, Zhu B, et al. (2018) CuInS2 sensitized TiO2 hybrid nanofibers for improved photocatalytic CO2 reduction. Appl Catal B-Environ 230: 194–202. doi: 10.1016/j.apcatb.2018.02.042
|
| [16] |
Camarillo R, Toston S, Martinez F, et al. (2017) Preparation of TiO2-based catalysts with supercritical fluid technology: characterization and photocatalytic activity in CO2 reduction. J Chem Technol Biot 92: 1710–1720. doi: 10.1002/jctb.5169
|
| [17] | Wei Y, Jiao J, Zhao Z, et al. (2015) 3D ordered macroporous TiO2-supported Pt@CdS core-shell nanoparticles: design, synthesis and efficient photocatalytic conversion of CO2 with water to methane. J Mater Chem A 20: 11074–11085. |
| [18] |
Zhao Y, Wei Y, Wu X, et al. (2018) Graphene-wrapped Pt/TiO2 photocatalysts with enhanced photogenerated charges separation and reactant adsorption for high selective photoreduction of CO2 to CH4. Appl Catal B-Environ 226: 360–372. doi: 10.1016/j.apcatb.2017.12.071
|
| [19] |
Toston S, Camarillo R, Martinez F, et al. (2017) Supercritical synthesis of platinum-modified titanium dioxide for solar fuel production from carbon dioxide. Chinese J Catal 38: 636–650. doi: 10.1016/S1872-2067(17)62766-9
|
| [20] |
Jiao J, Wei Y, Zhao Y, et al. (2017) AuPd/3DOM-TiO2 catalysts for photocatalyic reduction of CO2: high efficient separation of photogenerated charge carriers. Appl Catal B-Environ 209: 228–239. doi: 10.1016/j.apcatb.2017.02.076
|
| [21] |
Camrillo R, Toston S, Martinez F, et al. (2017) Enhancing the photocatalytic reduction of CO2 through engineering of catalysts with high pressure technology: Pd/TiO2 photocatalysts. J Supercrit Fluid 123: 18–27. doi: 10.1016/j.supflu.2016.12.010
|
| [22] |
Chen W, Wang Y, Shangguan W (2019) Metal (oxide) modified (M = Pd, Ag, Au and Cu) H2SrTa2O7 for photocatalytic CO2 reduction with H2O: the effect of cocatalysts on promoting activity toward CO and H2 evolution. Int J Hydrogen Energ 44: 4123–4132. doi: 10.1016/j.ijhydene.2018.12.166
|
| [23] |
Yu Y, Lan Z, Guo L, et al. (2018) Synergistic effects of Zn and Pd species in TiO2 towards efficient photo-reduction of CO2 into CH4. New J Chem 42: 483–488. doi: 10.1039/C7NJ03305B
|
| [24] |
Singhal N, Kumar U (2017) Noble metal modified TiO2: selective photoreduction of CO2 to hydrocarbons. Mol Catal 439: 91–99. doi: 10.1016/j.mcat.2017.06.031
|
| [25] |
Yui T, Kan A, Saitoh C, et al. (2011) Photochemical reduction of CO2 using TiO2: effects of organic adsorbates on TiO2 and deposition of Pd onto TiO2. ACS Appl Mater Inter 3: 2594–2600. doi: 10.1021/am200425y
|
| [26] | Li N, Liu M, Yang B, et al. (2017) Enhanced photocatalytic performance toward CO2 hydrogenation over nanosized TiO2-loaded Pd under UV irradiation. J Phys Chem 121: 2923–2932. |
| [27] |
Goren Z, Willner I, Nelson AJ, et al. (1990) Selective photoreduction of CO2/HCO3− to formate by aqueous suspensions and colloids of Pd-TiO2. J Phys Chem 94: 3784–3790. doi: 10.1021/j100372a080
|
| [28] |
Tseng IH, Chang WC, Wu JCS (2002) Photoreduction of CO2 using sol-gel derived titania and titania-supported copper catalysts. Appl Catal B-Environ 37: 37–38. doi: 10.1016/S0926-3373(01)00322-8
|
| [29] |
Nishimura A, Sugiura N, Fujita M, et al. (2007) Influence of preparation conditions of coated TiO2 film on CO2 reforming performance. Kagaku Kogaku Ronbun 33: 146–153. doi: 10.1252/kakoronbunshu.33.146
|
| [30] |
Izumi Y (2013) Recent advances in the photocatalytic conversion of carbon dioxide to fuels with water and/or hydrogen using solar energy and beyond. Coordin Chem Rev 257: 171–186. doi: 10.1016/j.ccr.2012.04.018
|
| [31] |
Lo CC, Hung CH, Yuan CS, et al. (2007) Photoreduction of carbon dioxide with H2 and H2O over TiO2 and ZrO2 in a circulated photocatalytic reactor. Sol Energ Mat Sol C 91: 1765–1774. doi: 10.1016/j.solmat.2007.06.003
|
| [32] |
Mahmodi G, Sharifnia S, Madani M, et al. (2013) Photoreduction of carbon dioxide in the presence of H2, H2O and CH4 over TiO2 and ZnO photocatalysts. Sol Energy 97: 186–194. doi: 10.1016/j.solener.2013.08.027
|
| [33] |
Jensen J, Mikkelsen M, Krebs FC (2011) Flexible substrates as basis for photocatalytic reduction of carbon dioxide. Sol Energ Mat Sol C 95: 2949–2958. doi: 10.1016/j.solmat.2011.06.040
|
| [34] | Nishimura A, Ishida N, Tatematsu D, et al. (2017) Effect of Fe loading condition and reductants on CO2 reduction performance with Fe/TiO2 photocatalyst. Int J Photoenergy 2017: 1625274. |
| [35] | Nishimura A, Tatematsu D, Toyoda R, et al. (2019) Effect of Overlapping Layout of Fe/TiO2 on CO2 Reduction with H2 and H2O. MOJ Sol Photoenergy Syst 3: 1–8. |
| [36] | Nishimura A, Toyoda R, Tatematsu D, et al. (2018) Effect of Molar Ratio of H2 and H2O on CO2 Reduction Characteristics of Overlapped Cu/TiO2. Int J Eng Sci Invent 7: 18–28. |
| [37] | Japan Society of Mechanical Engineering (1993) Heat Transfer Hand Book, 1 Eds, Tokyo: Maruzen. |
| [38] |
Wang D, Zhou WL, McCaughy BF, et al. (2003) Electrodeposition of metallic nanowire thin films using mesoporous silica templates. Adv Mater 15: 130–133. doi: 10.1002/adma.200390025
|
| [39] |
Koci K, Matejova L, Reli M, et al. (2014) Sol-gel derived Pd supported TiO2-ZrO2 and TiO2 photocatalysts; their examination in photocatalytic reduction of carbon dioxide. Catal Today 230: 20–26. doi: 10.1016/j.cattod.2013.10.002
|
| [40] |
Wei Y, Wu X, Zhao Y, et al. (2018) Efficient photocatalysts of TiO2 nanocrystals-supported PtRu alloy nanoparticles for CO2 reduction with H2O: synergistic effect of Pt-Ru. Appl Catal B-Environ 236: 445–457. doi: 10.1016/j.apcatb.2018.05.043
|
| [41] |
Zhang R, Huang Z, Li C, et al. (2019) Monolithic g-C3N4/reduced graphene oxide aerogel with in situ embedding of Pd nanoparticles for hydrogenation of CO2 to CH4. Appl Surf Sci 475: 953–960. doi: 10.1016/j.apsusc.2019.01.050
|
| [42] |
Kulandaivalu T, Rashid SA, Sabli N, et al. (2019) Visible light assisted photocatalytic reduction of CO2 to ethane using CQDs/Cu2O nanocomposite photocatalyst. Diam Relat Mater 91: 64–73. doi: 10.1016/j.diamond.2018.11.002
|
| [43] |
Zhu Z, Huang WR, Chen CY, et al. (2018) Preparation of Pd-Au/TiO2-WO3 to enhance photoreduction of CO2 to CH4 and CO. J CO2 Util 28: 247–254. doi: 10.1016/j.jcou.2018.10.006
|
| [44] |
Tasbihi M, Fresno F, Simon U, et al. (2018) On the selectivity of CO2 photoreduction towards CH4 using Pt/TiO2 catalysts supported on mesoporous silica. Appl Catal B-Environ 239: 68–76. doi: 10.1016/j.apcatb.2018.08.003
|
| [45] |
Wei Y, Wu X, Zhao Y, et al. (2018) Efficient photocatalysts of TiO2 nanocrystals-supported PtRu alloy nanoparticles for CO2 reduction with H2O: synergistic effect of Pt-Ru. Appl Catal B-Environ 236: 445–457. doi: 10.1016/j.apcatb.2018.05.043
|
| [46] |
Hashemizadeh I, Golovko VB, Choi J, et al. (2018) Photocatalytc reduction of CO2 to hydrocarbons using bio-templated porous TiO2 architectures under UV and visible light. Chem Eng J 347: 64–73. doi: 10.1016/j.cej.2018.04.094
|
| [47] |
Hoque MA, Guzman MJ (2018) Photocatalytic activity: experimental features to report in heterogeneous photocatalysis. Materials 11: 1990. doi: 10.3390/ma11101990
|
| [48] | Nishimura A, Okano Y, Hirota M, et al. (2011) Effect of preparation condition of TiO2 film and experimental condition on CO2 reduction performance of TiO2 photocatalyst membrane reactor. Int J Photoenergy 2011: 305650. |
| [49] |
Tahir M, Amin NS (2013) Advances in visible light responsive titanium oxide-based photocatalysts for CO2 conversion to hydrocarbon fuels. Energ Convers Manage 76: 194–214. doi: 10.1016/j.enconman.2013.07.046
|
| [50] |
Nagaveni K, Hedge MS, Madras G (2004) Structure and photocatalytic activity of Ti1−xMxO2±d (M = W, V, Ce, Zr, and Cu) synthesized by solution combustion method. J Phys Chem B 108: 20204–20212. doi: 10.1021/jp047917v
|
| [51] | Li L, Gao F, Zhao H, et al. (2013) Tailoring Cu valence and oxygen vacancy in Cu/TiO2 catalysts for enhanced CO2 photoreduction efficiency. Appl Catal B-Environ 134–135: 349–358. |
| [52] |
Song G, Xin F, Chen J, et al. (2014) Photocatalytic reduction of CO2 in cyclohexanol on CdS-TiO2 heterostructured photocatalyst. Appl Catal A-Gen 473: 90–95. doi: 10.1016/j.apcata.2013.12.035
|
| [53] |
Song G, Xin F, Yin X (2015) Photocatalytic reduction of carbon dioxide over ZnFe2O4/TiO2 nanobelts heterostructure in cyclohexanol. J Colloid Interf Sci 442: 60–66. doi: 10.1016/j.jcis.2014.11.039
|

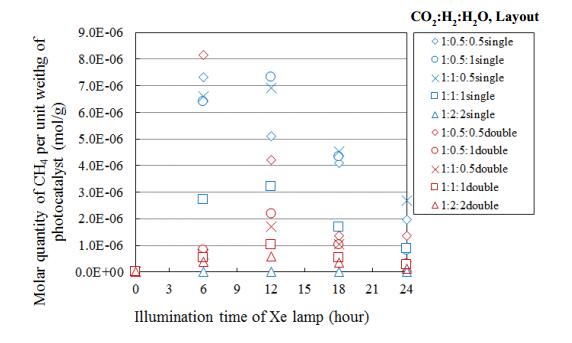
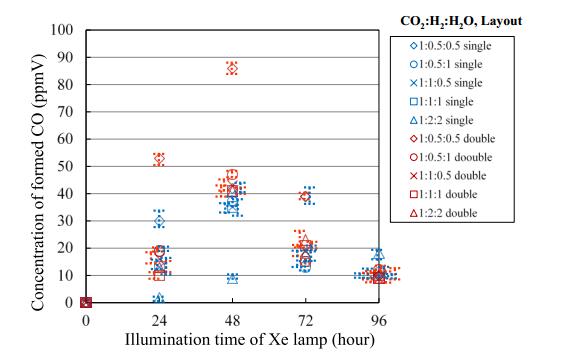
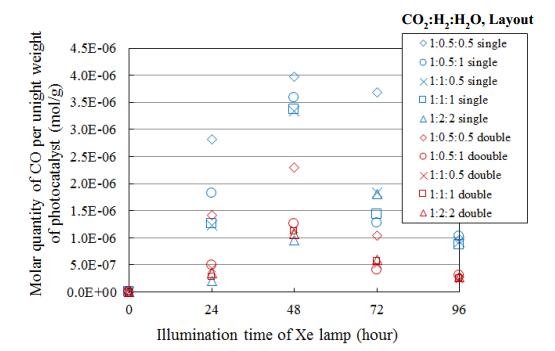
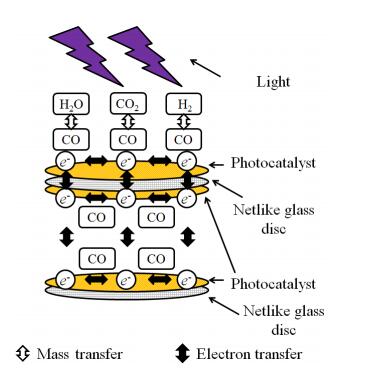
Figures(15) / Tables(4)

Akira Nishimura, Tadaki Inoue, Yoshito Sakakibara, Masafumi Hirota, Akira Koshio, Fumio Kokai, Eric Hu. Optimum molar ratio of H2 and H2O to reduce CO2 using Pd/TiO2[J]. AIMS Materials Science, 2019, 6(4): 464-483. doi: 10.3934/matersci.2019.4.464







 DownLoad:
DownLoad: