Citation: Carlo Danieli, Bertin Many Manda, Thudiyangal Mithun, Charalampos Skokos. Computational efficiency of numerical integration methods for the tangent dynamics of many-body Hamiltonian systems in one and two spatial dimensions[J]. Mathematics in Engineering, 2019, 1(3): 447-488. doi: 10.3934/mine.2019.3.447

| [1] | Fermi E, Pasta P, Ulam S, et al.(1955) Studies of the nonlinear problems, Los Alamos Report LA-1940. |
| [2] |
Ford J (1992) The Fermi-Pasta-Ulam problem: Paradox turns discovery. Phys Rep 213: 271–310. doi: 10.1016/0370-1573(92)90116-H

|
| [3] |
Campbell DK, Rosenau P, Zaslavsky GM (2005) Introduction: The Fermi-Pasta-Ulam problem: The first fifty years. Chaos 15: 015101. doi: 10.1063/1.1889345
|
| [4] |
Lepri S, Livi R, Politi A (2003) Universality of anomalous one-dimensional heat conductivity. Phys Rev E 68: 067102. doi: 10.1103/PhysRevE.68.067102
|
| [5] |
Lepri S, Livi R, Politi A (2005) Studies of thermal conductivity in Fermi-Pasta-Ulam-like lattices. Chaos 15: 015118. doi: 10.1063/1.1854281
|
| [6] |
Antonopoulos C, Bountis T, Skokos Ch (2006) Chaotic dynamics of N-degree of freedom Hamiltonian systems. Int J Bifurcation Chaos 16: 1777–1793. doi: 10.1142/S0218127406015672
|
| [7] | Zabusky NJ, Kruskal MD (1965) Interaction of "solitons" in a collisionless plasma and the recurrence of initial states. Phys Rev Lett 15: 240–243. |
| [8] | Zabusky NJ, Deem GS (1967) Dynamics of nonlinear lattices I. Localized optical excitations, acoustic radiation, and strong nonlinear behavior. J Comput Phys 2: 126–153. |
| [9] | Izrailev FM, Chirikov BV (1966) Statistical properties of a nonlinear string. Sov Phys Dokl 11: 30pages. |
| [10] |
Paleari S, Penati T (2008) Numerical methods and results in the FPU problem. Lect Notes Phys 728: 239–282. doi: 10.1007/978-3-540-72995-2_7
|
| [11] |
Anderson PW (1958) Absence of diffusion in certain random lattices. Phys Rev 109: 1492–1505. doi: 10.1103/PhysRev.109.1492
|
| [12] |
Abraham E, Anderson PW, Licciardello DC, et al. (1979) Scaling theory of localization: Absence of quantum diffusion in two dimensions. Phys Rev Lett 42: 673–676. doi: 10.1103/PhysRevLett.42.673
|
| [13] |
Shepelyansky DL (1993) Delocalization of quantum chaos by weak nonlinearity. Phys Rev Lett 70: 1787–1790. doi: 10.1103/PhysRevLett.70.1787
|
| [14] |
Molina MI (1998) Transport of localized and extended excitations in a nonlinear Anderson model. Phys Rev B 58: 12547–12550. doi: 10.1103/PhysRevB.58.12547
|
| [15] |
Clément D, Varon AF, Hugbart M, et al. (2005) Suppression of transport of an interacting elongated Bose-Einstein condensate in a random potential. Phys Rev Lett 95: 170409. doi: 10.1103/PhysRevLett.95.170409
|
| [16] |
Fort C, Fallani L, Guarrera V, et al. (2005) Effect of optical disorder and single defects on the expansion of a Bose-Einstein condensate in a one-dimensional waveguide. Phys Rev Lett 95: 170410. doi: 10.1103/PhysRevLett.95.170410
|
| [17] |
Schwartz T, Bartal G, Fishman S, et al. (2007) Transport and Anderson localization in disordered two-dimensional photonic lattices. Nature 446: 52–55. doi: 10.1038/nature05623
|
| [18] |
Lahini Y, Avidan A, Pozzi F, et al. (2008) Anderson localization and nonlinearity in one-dimensional disordered photonic lattices. Phys Rev Lett 100: 013906. doi: 10.1103/PhysRevLett.100.013906
|
| [19] |
Billy J, Josse V, Zuo Z, et al. (2008) Direct observation of Anderson localization of matter waves in a controlled disorder. Nature 453: 891–894. doi: 10.1038/nature07000
|
| [20] |
Roati JG, D'Errico C, Fallani L, et al. (2008) Anderson localization of a non-interacting Bose–Einstein condensate. Nature 453: 895–898. doi: 10.1038/nature07071
|
| [21] |
Fallani L, Fort C, Inguscio M (2008) Bose-Einstein condensates in disordered potentials. Adv At Mol Opt Phys 56: 119–160. doi: 10.1016/S1049-250X(08)00012-8
|
| [22] |
Flach S, Krimer DO, Skokos Ch (2009) Universal spreading of wave packets in disordered nonlinear systems. Phys Rev Lett 102: 024101. doi: 10.1103/PhysRevLett.102.024101
|
| [23] |
Vicencio RA, Flach S (2009) Control of wave packet spreading in nonlinear finite disordered lattices. Phys Rev E 79: 016217. doi: 10.1103/PhysRevE.79.016217
|
| [24] |
Skokos Ch, Krimer DO, Komineas S, et al. (2009) Delocalization of wave packets in disordered nonlinear chains. Phys Rev E 79: 056211. doi: 10.1103/PhysRevE.79.056211
|
| [25] |
Skokos Ch, Flach S (2010) Spreading of wave packets in disordered systems with tunable nonlinearity. Phys Rev E 82: 016208. doi: 10.1103/PhysRevE.82.016208
|
| [26] |
Laptyeva TV, Bodyfelt JD, Krimer DO, et al. (2010) The crossover from strong to weak chaos for nonlinear waves in disordered systems. EPL 91: 30001. doi: 10.1209/0295-5075/91/30001
|
| [27] |
Modugno G (2010) Anderson localization in Bose-Einstein condensates. Rep Prog Phys 73: 102401. doi: 10.1088/0034-4885/73/10/102401
|
| [28] | Bodyfelt JD, Laptyeva TV, Gligoric G, et al. (2011) Wave interactions in localizing media - a coin with many faces, Int J Bifurcat Chaos 21: 2107. |
| [29] |
Bodyfelt JD, Laptyeva TV, Skokos Ch, et al. (2011) Nonlinear waves in disordered chains: Probing the limits of chaos and spreading. Phys Rev E 84: 016205. doi: 10.1103/PhysRevE.84.016205
|
| [30] |
Laptyeva TV, Bodyfelt JD, Flach S (2012) Subdiffusion of nonlinear waves in two-dimensional disordered lattices. EPL 98: 60002. doi: 10.1209/0295-5075/98/60002
|
| [31] |
Pikovsky AS, Shepelyansky DL (2008) Destruction of Anderson localization by a weak nonlinearity. Phys Rev Lett 100: 094101. doi: 10.1103/PhysRevLett.100.094101
|
| [32] |
Skokos Ch, Gkolias I, Flach S (2013) Nonequilibrium chaos of disordered nonlinear waves. Phys Rev Lett 111: 064101. doi: 10.1103/PhysRevLett.111.064101
|
| [33] |
Senyangue B, Many Manda B, Skokos Ch (2018) Characteristics of chaos evolution in one- dimensional disordered nonlinear lattices. Phys Rev E 98: 052229. doi: 10.1103/PhysRevE.98.052229
|
| [34] | Kati Y, Yu X, Flach S (2019) Density resolved wave packet spreading in disordered Gross-Pitaevskii lattices. In preparation. |
| [35] | Benettin G, Galgani L, Giorgilli A, et al. (1980) Lyapunov characteristic exponents for smooth dynamical systems and for Hamiltonian systems; a method for computing all of them. Part 2: Numerical application. Meccanica 15: 21–30. |
| [36] |
Benettin G, Galgani L, Giorgilli A, et al. (1980) Lyapunov characteristic exponents for smooth dynamical systems and for Hamiltonian systems; a method for computing all of them. Part 1: Theory. Meccanica 15: 9–20. doi: 10.1007/BF02128236
|
| [37] |
Skokos Ch (2010) The Lyapunov characteristic exponents and their computation. Lect Notes Phys 790: 63–135. doi: 10.1007/978-3-642-04458-8_2
|
| [38] |
Mulansky M, Ahnert K, Pikovsky A, et al. (2009) Dynamical thermalization of disordered nonlinear lattices. Phys Rev E 80: 056212. doi: 10.1103/PhysRevE.80.056212
|
| [39] |
Sales MO, Dias WS, Neto AR, et al. (2018) Sub-diffusive spreading and anomalous localization in a 2D Anderson model with off-diagonal nonlinearity. Solid State Commun 270: 6–11. doi: 10.1016/j.ssc.2017.11.001
|
| [40] | Hairer E, Lubich C, Wanner G (2002) Geometric Numerical Integration. Structure-Preservings Algorithms for Ordinary Differential Equations. Springer-Verlag Berlin Heidelberg. Vol 31. |
| [41] | McLachlan RI, Quispel GRW (2002) Splitting methods. Acta Numer 11: 341–434. |
| [42] |
McLachlan RI, Quispel GRW (2006) Geometric integrators for ODEs. J Phys A Math Gen 39: 5251–5285. doi: 10.1088/0305-4470/39/19/S01
|
| [43] | Forest E (2006) Geometric integration for particle accelerators. J Phys A Math Gen 39: 5351-5377. |
| [44] | Blanes S, Casas F, Murua A (2008) Splitting and composition methods in the numerical integration of differential equations. Bol Soc Esp Mat Apl 45: 89–145. |
| [45] |
Benettin G, Ponno A (2011) On the numerical integration of FPU-like systems. Phys D 240: 568–573. doi: 10.1016/j.physd.2010.11.008
|
| [46] |
Antonopoulos Ch, Bountis T, Skokos Ch, et al. (2014) Complex statistics and diffusion in nonlinear disordered particle chains. Chaos 24: 024405. doi: 10.1063/1.4871477
|
| [47] |
Antonopoulos Ch, Skokos Ch, Bountis T, et al. (2017) Analyzing chaos in higher order disordered quartic-sextic Klein-Gordon lattices using q-statistics. Chaos Solitons Fractals 104: 129–134. doi: 10.1016/j.chaos.2017.08.005
|
| [48] |
Tieleman O, Skokos Ch, Lazarides A (2014) Chaoticity without thermalisation in disordered lattices. EPL 105: 20001. doi: 10.1209/0295-5075/105/20001
|
| [49] |
Skokos Ch, Gerlach E, Bodyfelt JD, et al. (2014) High order three part split symplectic integrators: Efficient techniques for the long time simulation of the disordered discrete non linear Schrödinger equation. Phys Lett A 378: 1809–1815. doi: 10.1016/j.physleta.2014.04.050
|
| [50] |
Gerlach E, Meichsner J, Skokos C (2016) On the symplectic integration of the discrete nonlinear Schrödinger equation with disorder. Eur Phys J Spec Top 225: 1103–1114. doi: 10.1140/epjst/e2016-02657-0
|
| [51] |
Danieli C, Campbell DK, Flach S (2017) Intermittent many-body dynamics at equilibrium. Phys Rev E 95: 060202. doi: 10.1103/PhysRevE.95.060202
|
| [52] |
Thudiyangal M, Kati Y, Danieli C, et al. (2018) Weakly nonergodic dynamics in the Gross-Pitaevskii lattice. Phys Rev Lett 120: 184101. doi: 10.1103/PhysRevLett.120.184101
|
| [53] |
Thudiyangal M, Danieli C, Kati Y, et al. (2019) Dynamical glass phase and ergodization times in Josephson junction chains. Phys Rev Lett 122: 054102. doi: 10.1103/PhysRevLett.122.054102
|
| [54] | Danieli C, Thudiyangal M, Kati Y, et al. (2018) Dynamical glass in weakly non-integrable many-body systems. arXiv:1811.10832. |
| [55] |
Laskar J, Robutel P (2001) High order symplectic integrators for perturbed Hamiltonian systems. Celest Mech Dyn Astr 80: 39–62. doi: 10.1023/A:1012098603882
|
| [56] |
Senyange B, Skokos Ch (2018) Computational efficiency of symplectic integration schemes: Application to multidimensional disordered Klein-Gordon lattices. Eur Phys J Spec Top 227: 625–643. doi: 10.1140/epjst/e2018-00131-2
|
| [57] |
Blanes S, Casas F, Farrés A, et al. (2013) New families of symplectic splitting methods for numerical integration in dynamical astronomy. Appl Numer Math 68: 58–72. doi: 10.1016/j.apnum.2013.01.003
|
| [58] | Skokos Ch, Gerlach E (2010) Numerical integration of variational equations. Phys Rev E 82: 036704. |
| [59] | Gerlach E, Skokos Ch (2011) Comparing the efficiency of numerical techniques for the integration of variational equations: Dynamical systems, differential equations and applications, Discrete & Continuous Dynamical Systems-Supplement 2011, Dedicated to the 8th AIMS Conference, 475–484. |
| [60] |
Gerlach E, Eggl S, Skokos Ch (2012) Efficient integration of the variational equations of multidimensional Hamiltonian systems: Application to the Fermi-Pasta-Ulam lattice. Int J Bifurcation Chaos 22: 1250216. doi: 10.1142/S0218127412502161
|
| [61] |
Carati A, Ponno A (2018) Chopping time of the FPU α-model. J Stat Phys 170: 883–894. doi: 10.1007/s10955-018-1962-8
|
| [62] |
Flach S, Ivanchenko MV, Kanakov OI (2005) q-Breathers and the Fermi-Pasta-Ulam problem. Phys Rev Lett 95: 064102. doi: 10.1103/PhysRevLett.95.064102
|
| [63] |
Flach S, Ivanchenko MV, Kanakov OI, et al. (2008) Periodic orbits, localization in normal mode space, and the Fermi-Pasta-Ulam problem. Am J Phys 76: 453. doi: 10.1119/1.2820396
|
| [64] |
Flach S, Ponno A (2008) The Fermi-Pasta-Ulam problem: periodic orbits, normal forms and resonance overlap criteria. Phys D 237: 908–917. doi: 10.1016/j.physd.2007.11.017
|
| [65] |
Garcia-Mata I, Shepelyansky DL (2009) Delocalization induced by nonlinearity in systems with disorder. Phys Rev E 79: 026205. doi: 10.1103/PhysRevE.79.026205
|
| [66] | Hairer E, Nørsett SP, Wanner G (1993) Solving Ordinary Differential Equations I: Nonstiff Problems, 2Ed., Berlin: Springer, Vol. 14. |
| [67] | Barrio R (2005) Performance of the Taylor series method for ODEs/DAEs. Appl Math Comput 163: 525–545. |
| [68] | Abad A, Barrio R, Blesa F, et al. (2012) Algorithm 924: TIDES, a Taylor series integrator for differential equations. ACM Math Software 39: 5. |
| [69] | Taylor series Integrator for Differential Equations. Available from: https://sourceforge.net/projects/tidesodes/. |
| [70] | Gröbner W (1967) Die Lie-Reihen und ihre Anwendungen. Berlin: Deutscher Verlag der Wissenschaften. |
| [71] | Blanes S, Casas F (2016) A Concise Introduction to Geometric Numerical Integration. In series of Monographs and Research Notes in Mathematics, Chapman and Hall/CRC. |
| [72] | Hanslmeier A, Dvorak R (1984) Numerical integration with Lie series. Astron Astrophys 132: 203–207. |
| [73] |
Eggl S, Dvorak R (2010) An introduction to common numerical integration codes used in dynamical astronomy. Lect Notes Phys 790: 431–480. doi: 10.1007/978-3-642-04458-8_9
|
| [74] | Fortran and Matlab Codes. Available from: http://www.unige.ch/˜hairer/software.html . |
| [75] | Boreux J, Carletti T, Hubaux C (2010) High order explicit symplectic integrators for the Discrete Non Linear Schrödinger equation. Report naXys 09, arXiv:1012.3242. |
| [76] |
Laskar J (2003) Chaos in the solar system. Ann Henri Poincaré 4: 693–705. doi: 10.1007/s00023-003-0955-5
|
| [77] | Leimkuhler B, Reich S (2004) Simulating Hamiltonian Dynamics. UK: Cambridge University Press, Vol. 14. |
| [78] | Lasagni FM (1988) Canonical Runge-Kutta methods. ZAMP 39: 952–953. |
| [79] |
Sanz-Serna JM (1988) Runge-Kutta schemes for Hamiltonian systems. BIT Numer Math 28: 877–883. doi: 10.1007/BF01954907
|
| [80] |
Yoshida H (1990) Construction of higher order symplectic integrator. Phys Lett A 150: 262–268. doi: 10.1016/0375-9601(90)90092-3
|
| [81] |
Suzuki M (1990) Fractal decomposition of exponential operators with applications to many-body theories and Monte Carlo simulations. Phys Lett A 146: 319–323. doi: 10.1016/0375-9601(90)90962-N
|
| [82] |
Yoshida H (1993) Recent progress in the theory and application of symplectic integrators. Celest Mech Dyn Astr 56: 27–43. doi: 10.1007/BF00699717
|
| [83] |
Ruth RD (1983) A canonical integration technique. IEEE Trans Nucl Sci 30: 2669–2671. doi: 10.1109/TNS.1983.4332919
|
| [84] |
McLachlan RI (1995) Composition methods in the presence of small parameters. BIT Numer Math 35: 258–268. doi: 10.1007/BF01737165
|
| [85] |
Farrés A, Laskar J, Blanes S, et al. (2013) High precision symplectic integrators for the solar system. Celest Mech Dyn Astr 116: 141–174. doi: 10.1007/s10569-013-9479-6
|
| [86] | Iserles A, Quispel GRW (2018) Why geometric numerical integration? In: Ebrahimi-Fard, K., Barbero Liñán, M. Editors, Discrete Mechanics, Geometric Integration and Lie-Butcher Series, Springer, Cham. |
| [87] |
Forest E, Ruth RD (1990) Fourth-order symplectic integration. Phys D 43: 105–117. doi: 10.1016/0167-2789(90)90019-L
|
| [88] |
Kahan W, Li RC (1997) Composition constants for raising the orders of unconventional schemes for ordinary differential equations. Math Comput Am Math Soc 66: 1089–1099. doi: 10.1090/S0025-5718-97-00873-9
|
| [89] |
Sofroniou M, Spaletta G (2005) Derivation of symmetric composition constants for symmetric integrators. Optim Method Soft 20: 597–613. doi: 10.1080/10556780500140664
|
| [90] | Blanes S, Moan PC (2001) Practical symplectic partitioned Runge-Kutta and Runge-Kutta-Nyström methods. J Comput Appl Math 142: 313–330. |
| [91] |
Hillebrand M, Kalosakas G, Schwellnus A, et al. (2019) Heterogeneity and chaos in the Peyrard- Bishop-Dauxois DNA model. Phys Rev E 99: 022213. doi: 10.1103/PhysRevE.99.022213
|
| [92] |
Laskar J, Vaillant T (2019) Dedicated symplectic integrators for rotation motions. Celest Mech Dyn Astr 131: 15. doi: 10.1007/s10569-019-9886-4
|
| [93] | Koseleff PV (1996) Exhaustive search of symplectic integrators using computer algebra. Integr Algorithms Classical Mech 10: 103–120. |
| [94] |
Benettin G, Galgani L, Giorgilli A, et al. (1976) Kolmogorov entropy and numerical experiments. Phys Rev A 14: 2338–2345. doi: 10.1103/PhysRevA.14.2338
|
| [95] | All simulations were performed on the IBS-PCS cluster, which uses Intel(R) Xeon(R) E5-2620 v3 processors. All codes were written in Fortran90 language and were compiled by using the gfortran compiler ( https://gcc.gnu.org/ ) with O3 optimization flag. No advanced vectorization mode has been implemented. |
| [96] | All simulations were performed on a workstation using 3.00 GHz Intel Xeon E5-2623 processors. All codes were written in Fortran90 language and were compiled by using the gfortran compiler ( https://gcc.gnu.org/ ) with O3 optimization flag. No advanced vectorization mode has been implemented. |
| [97] |
Rasmussen K, Cretegny T, Kevrekidis PG, et al. (2000) Statistical mechanics of a discrete nonlinear system. Phys Rev Lett 84: 3740–3743. doi: 10.1103/PhysRevLett.84.3740
|
| [98] |
Skokos Ch, Manos T (2016) The Smaller (SALI) and the Generalized (GALI) alignment indices: Efficient methods of chaos detection. Lect Notes Phys 915: 129–181. doi: 10.1007/978-3-662-48410-4_5
|
| [99] |
Achilleos V, Theocharis G, Skokos Ch (2016) Energy transport in one-dimensional disordered granular solids. Phys Rev E 93: 022903. doi: 10.1103/PhysRevE.93.022903
|
| [100] |
Livi R, Pettini M, Ruffo S, et al. (1987) Chaotic behavior in nonlinear Hamiltonian systems and equilibrium statistical mechanics. J Stat Phys 48: 539–559. doi: 10.1007/BF01019687
|
| [101] |
Binder P, Abraimov D, Ustinov AV, et al. (2000) Observation of breathers in Josephson ladders. Phys Rev Lett 84: 745–748. doi: 10.1103/PhysRevLett.84.745
|
| [102] |
Blackburn JA, Cirillo M, Grønbech-Jensen N (2016) A survey of classical and quantum interpretations of experiments on Josephson junctions at very low temperatures. Phys Rep 611: 1–34. doi: 10.1016/j.physrep.2015.10.010
|
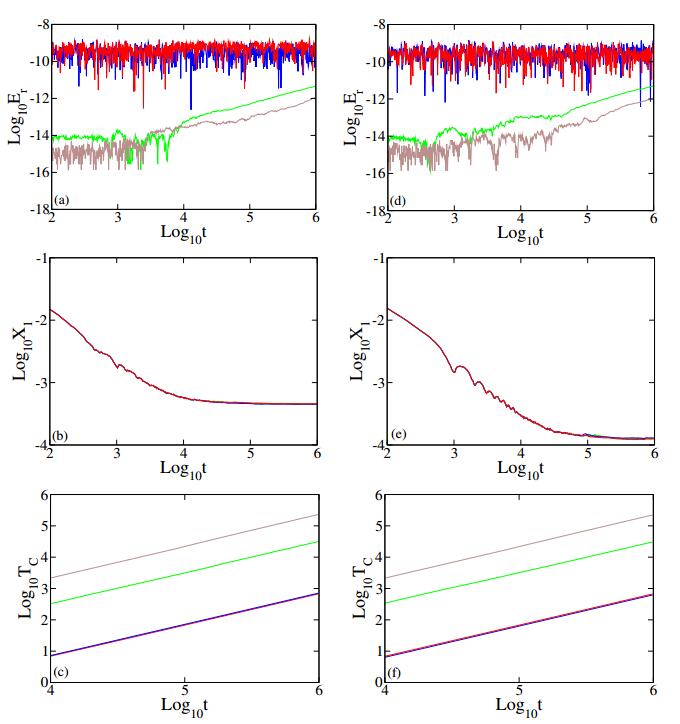
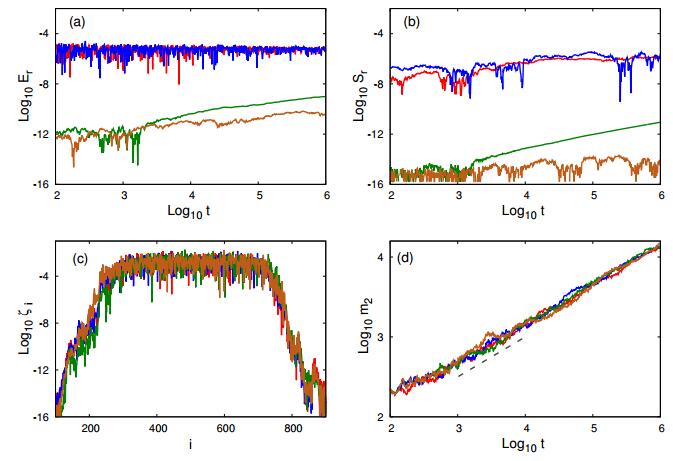
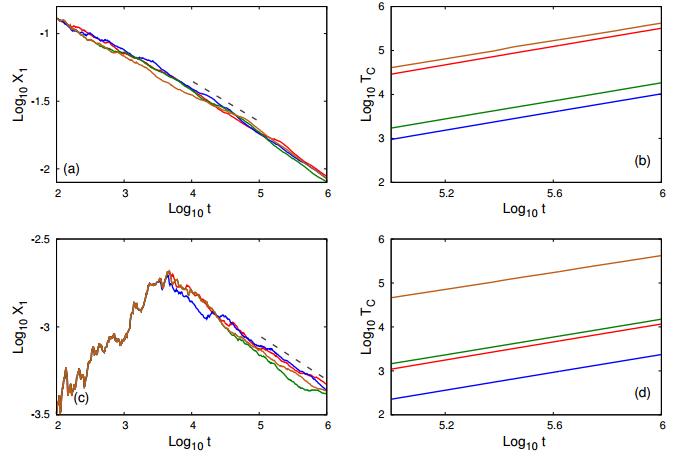
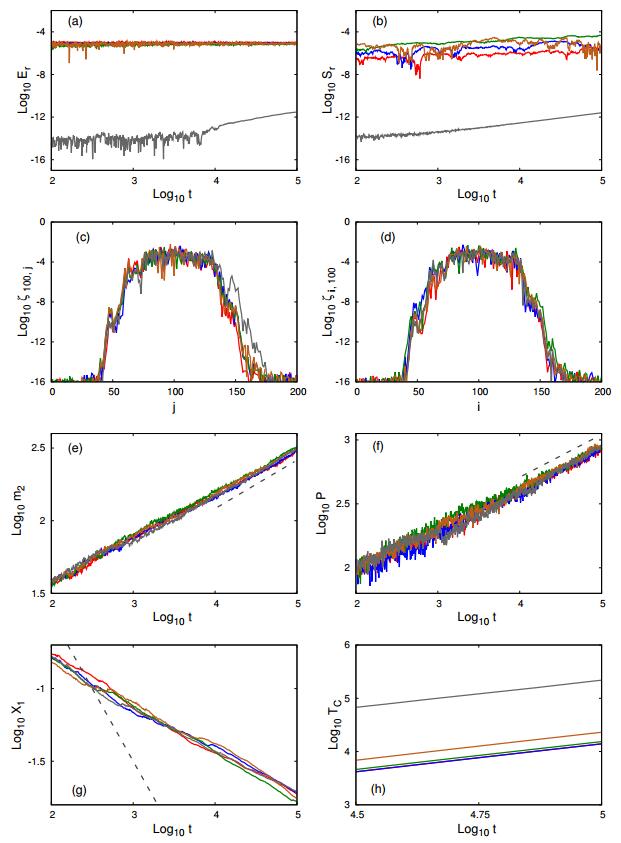
Figures(4) / Tables(6)

Carlo Danieli, Bertin Many Manda, Thudiyangal Mithun, Charalampos Skokos. Computational efficiency of numerical integration methods for the tangent dynamics of many-body Hamiltonian systems in one and two spatial dimensions[J]. Mathematics in Engineering, 2019, 1(3): 447-488. doi: 10.3934/mine.2019.3.447







 DownLoad:
DownLoad: