Citation: Ranjit Kumar Upadhyay, Swati Mishra. Population dynamic consequences of fearful prey in a spatiotemporal predator-prey system[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 338-372. doi: 10.3934/mbe.2019017

| [1] | B.R. Anholt and E.E.Werner, Density-dependent consequences of induced behavior, in The ecology and evolution of inducible defenses (eds. R. Tollrian and C.D. Harvell), Princeton University Press, USA, 1999, 218–230. |
| [2] | Z. Barta, A. Liker and F. Mónus, The effects of predation risk on the use of social foraging tactics, Anim. Behav., 67 (2004), 301–308. |
| [3] | B.A. Belgrad and B.D. Griffen, Predator–prey interactions mediated by prey personality and predator hunting mode, Proc. R. Soc. B, 283 (2016), 20160408. |
| [4] | C. Boesch, The effects of leopard predation on grouping patterns in forest chimpanzees, Behaviour, 117 (1991), 220–241. |
| [5] | M.J. Childress and M.A. Lung, Predation risk, gender and the group size effect: does elk vigilance depend upon the behaviour of conspecifics?, Anim. Behav., 66 (2003), 389–398. |
| [6] | S.N. Chow and J.K. Hale, Methods of Bifurcation Theory, vol. 251, Springer-Verlag, New York, 1982. |
| [7] | S. Creel and D. Christianson, Relationships between direct predation and risk effects, Trends Ecol. Evol., 23 (2008), 194–201. |
| [8] | S. Creel, D. Christianson, S. Liley and J.A. Winnie Jr., Predation risk affects reproductive physiology and demography of elk, Science, 315 (2007), 960–960. |
| [9] | S. Creel, J. Winnie Jr., B. Maxwell, K. Hamlin and M. Creel, Elk alter habitat selection as an antipredator response to wolves, Ecology, 86 (2005), 3387–3397. |
| [10] | D. Fortin, H.L. Beyer, M.S. Boyce, D.W. Smith, T. Duchesne and J.S. Mao, Wolves influence elk movements: behavior shapes a trophic cascade in Yellowstone National Park, Ecology, 86 (2005), 1320–1330. |
| [11] | M.R. Garvie, Finite-Difference Schemes for Reaction–Diffusion Equations Modeling Predator– Prey Interactions in MATLAB, Bull. Math. Biol., 69 (2007), 931–956. |
| [12] | B.D. Hassard, N.D. Kazarinoff and Y.H. Wan, Theory and applications of Hopf bifurcation, vol. 41, CUP Archive, 1981. |
| [13] | M. Hebblewhite, C.A. White, C.G. Nietvelt, J.A. McKenzie, T.E. Hurd, J.M. Fryxell, S.E. Bayley and P.C. Paquet, Human activity mediates a trophic cascade caused by wolves, Ecology, 86 (2005), 2135–2144. |
| [14] | D.S. Hik, Does risk of predation influence population dynamics? Evidence from the cyclic decline of snowshoe hares, Wildl. Res., 22 (1995), 115–129. |
| [15] | S.B. Hsu and T.W. Huang, Global stability for a class of predator–prey systems, SIAM J. Appl. Math., 55 (1995), 763–783. |
| [16] | M.S. Khudr, O.Y. Buzhdygan, J.S. Petermann and S.Wurst, Fear of predation alters clone-specific performance in phloem-feeding prey, Sci. Rep., 7 (2017), 7695. |
| [17] | C.J. Krebs, Two complementary paradigms for analysing population dynamics, Philos. Trans. R. Soc. Lond. B. Biol. Sci., 357 (2002), 1211–1219. |
| [18] | S.L. Lima, Nonlethal effects in the ecology of predator-prey interactions, Bioscience, 48 (1998), 25–34. |
| [19] | S.L. Lima, Predators and the breeding bird: behavioral and reproductive flexibility under the risk of predation, Biol. Rev., 84 (2009), 485–513. |
| [20] | S.L. Lima and P.A. Bednekoff, Temporal variation in danger drives antipredator behavior: the predation risk allocation hypothesis, Am. Nat., 153 (1999), 649–659. |
| [21] | S.J. McCauley, L. Rowe and M.J. Fortin, The deadly effects of |
| [22] | J.P. Michaud, P.R.R. Barbosa, C.L. Bain and J.B. Torres, Extending the |
| [23] | E.H. Nelson, C.E. Matthews and J.A. Rosenheim, Predators reduce prey population growth by inducing changes in prey behavior, Ecology, 85 (2004), 1853–1858. |
| [24] | K.L. Pangle, S.D. Peacor and O.E. Johannsson, Large nonlethal effects of an invasive invertebrate predator on zooplankton population growth rate, Ecology, 88 (2007), 402–412. |
| [25] | M. Pascual, Diffusion-induced chaos in a spatial predator–prey system, in Proc. R. Soc. Lond. B, vol. 251, 1993, 1–7. |
| [26] | B.L. Peckarsky, C.A. Cowan, M.A. Penton and C. Anderson, Sublethal consequences of streamdwelling predatory stoneflies on mayfly growth and fecundity, Ecology, 74 (1993), 1836–1846. |
| [27] | E.L. Preisser, D.I. Bolnick and M.F. Benard, Scared to death? The effects of intimidation and consumption in predator–prey interactions, Ecology, 86 (2005), 501–509. |
| [28] | R.M. Sapolsky, Why Zebras Don't Get Ulcers, 3rd edn., Henry Holt and Company, New York, NY, 2004. |
| [29] | O.J. Schmitz, A.P. Beckerman and K.M. O'Brien, Behaviorally mediated trophic cascades: effects of predation risk on food web interactions, Ecology, 78 (1997), 1388–1399. |
| [30] | G. Sun, S. Sarwardi, P.J. Pal and M.S. Rahman, The spatial patterns through diffusion-driven instability in modified Leslie-Gower and Holling-type II predator-prey model, J. Biol. System., 18 (2010), 593–603. |
| [31] | J.T. Tanner, The stability and the intrinsic growth rates of prey and predator populations, Ecology, 56 (1975), 855–867. |
| [32] | M. Travers, M. Clinchy, L. Zanette, R. Boonstra and T.D. Williams, Indirect predator effects on clutch size and the cost of egg production, Ecol. Lett., 13 (2010), 980–988. |
| [33] | R.K. Upadhyay, V. Volpert and N.K. Thakur, Propagation of Turing patterns in a plankton model, J. Biol. Dynam., 6 (2012), 524–538. |
| [34] | R.K. Upadhyay and P. Roy, Disease spread and its effect on population dynamics in heterogeneous environment, Int. J. Bifurc. Chaos, 26 (2016), 1650004. |
| [35] | R.K. Upadhyay, P. Roy and J. Datta, Complex dynamics of ecological systems under nonlinear harvesting: Hopf bifurcation and Turing instability, Nonlinear Dyn., 79 (2015), 2251–2270. |
| [36] | C.Wang, L. Chang and H. Liu, Spatial patterns of a predator-prey system of Leslie type with time delay, PloS one, 11 (2016), e0150503. |
| [37] | X. Wang, L. Zanette and X. Zou, Modelling the fear effect in predator–prey interactions, J. Math. Biol., 73 (2016), 1179–1204. |
| [38] | X. Wang and X. Zou, Modeling the Fear Effect in Predator–Prey Interactions with Adaptive Avoidance of Predators, Bull. Math. Biol., 79 (2017), 1325–1359. |
| [39] | X. Wang and X. Zou, Pattern formation of a predator-prey model with the cost of anti–predator behaviors, Math. Biosci. Engineer., 15 (2018), 775–805. |
| [40] | J. Winnie Jr., D. Christianson, S. Creel and B. Maxwell, Elk decision-making rules are simplified in the presence of wolves, Behav. Ecol. Sociobiol., 61 (2006), 277–289. |
| [41] | J. Winnie Jr. and S. Creel, Sex-specific behavioural responses of elk to spatial and temporal variation in the threat of wolf predation, Anim. Behav., 73 (2007), 215–225. |
| [42] | L.Y. Zanette, A.F. White, M.C. Allen and M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. |
| [43] | J.F. Zhang, W.T. Li and X.P. Yan, Hopf bifurcations in a predator-prey diffusion system with Beddington-DeAngelis response, Acta Appl. Math., 115 (2011), 91–104. |
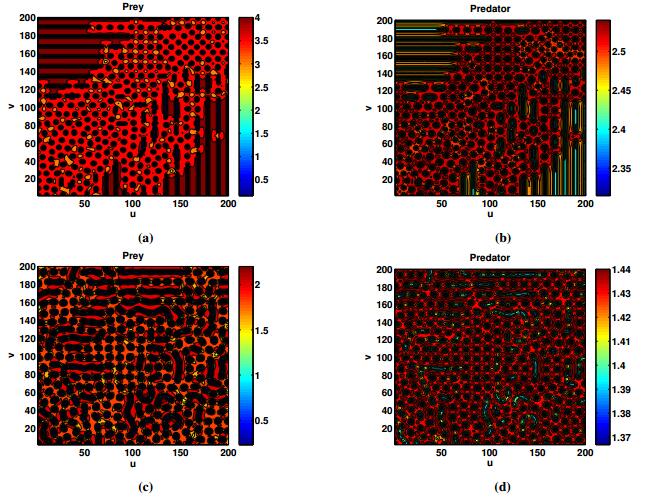
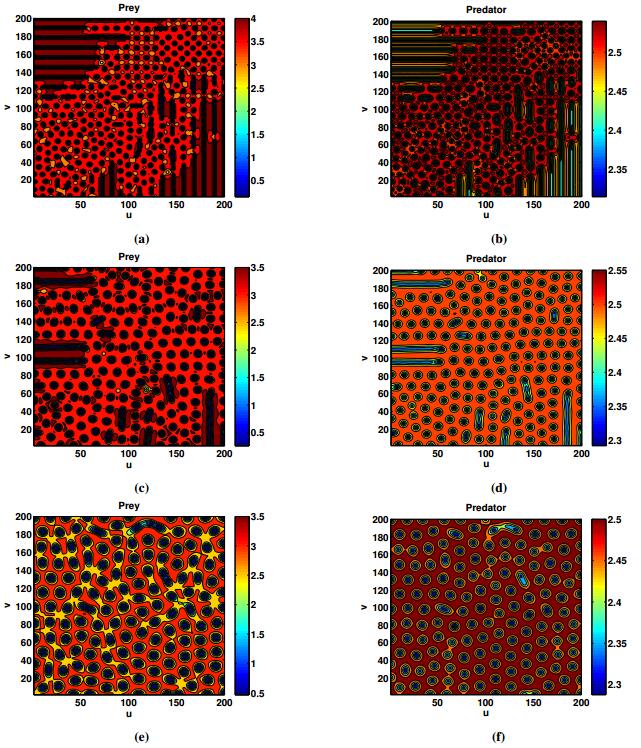
Figures(18) / Tables(2)

Ranjit Kumar Upadhyay, Swati Mishra. Population dynamic consequences of fearful prey in a spatiotemporal predator-prey system[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 338-372. doi: 10.3934/mbe.2019017







 DownLoad:
DownLoad: