Citation: Pranav Chintamani Joshi, Sugato Benerjee. Effects of glucocorticoids in depression: Role of astrocytes[J]. AIMS Neuroscience, 2018, 5(3): 200-210. doi: 10.3934/Neuroscience.2018.3.200

| [1] | Volterra A, Magistretti PJ, Haydon PG (2002) The tripartite synapse: Glia in synaptic transmission. New York: Oxford University Press. |
| [2] |
Pelvig DP, Pakkenberg H, Stark AK, et al. (2008) Neocortical glial cell numbers in human brains. Neurobiol Aging 29: 1754–1762. doi: 10.1016/j.neurobiolaging.2007.04.013

|
| [3] |
Ge WP, Jia JM (2016) Local production of astrocytes in the cerebral cortex. Neuroscience 323: 3–9. doi: 10.1016/j.neuroscience.2015.08.057
|
| [4] |
Hertz L (2008) Bioenergetics of cerebral ischemia: A cellular perspective. Neuropharmacology 55: 289–309. doi: 10.1016/j.neuropharm.2008.05.023
|
| [5] |
Bröer S, Brookes N (2001) Transfer of glutamine between astrocytes and neurons. J Neurochem 77: 705–719. doi: 10.1046/j.1471-4159.2001.00322.x
|
| [6] |
Halassa MM, Fellin T, Haydon PG (2007) The tripartite synapse: Roles for gliotransmission in health and disease. Trends Mol Med 13: 54–63. doi: 10.1016/j.molmed.2006.12.005
|
| [7] |
Nualart-Marti A, Solsona C, Fields RD (2013) Gap junction communication in myelinating glia. Biochim Biophys Acta 1828: 69–78. doi: 10.1016/j.bbamem.2012.01.024
|
| [8] | Cabezas R, Avila M, Gonzalez J, et al. (2014) Astrocytic modulation of blood brain barrier: Perspectives on Parkinson's disease. Front Cell Neurosci 8: 211. |
| [9] | Sapolsky RM, Romero LM, Munck AU (2000) How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev 21: 55–89. |
| [10] |
Chrousos GP (2009) Stress and disorders of the stress system. Nat Rev Endocrinol 5: 374–381. doi: 10.1038/nrendo.2009.106
|
| [11] |
De Kloet ER, Van Acker SA, Sibug RM, et al. (2000) Brain mineralocorticoid receptors and centrally regulated functions. Kidney Int 57: 1329–1336. doi: 10.1046/j.1523-1755.2000.00971.x
|
| [12] |
Carter BS, Meng F, Thompson RC (2012) Glucocorticoid treatment of astrocytes results in temporally dynamic transcriptome regulation and astrocyte-enriched mRNA changes in vitro. Physiol Genomics 44: 1188–1200. doi: 10.1152/physiolgenomics.00097.2012
|
| [13] |
Mcewen BS (2005) Glucocorticoids, depression, and mood disorders: Structural remodeling in the brain. Metabolism 54: 20–23. doi: 10.1016/j.metabol.2005.01.008
|
| [14] |
Green KN, Billings LM, Roozendaal B, et al. (2006) Glucocorticoids increase amyloid-beta and tau pathology in a mouse model of Alzheimer's disease. J Neurosci 26: 9047–9056. doi: 10.1523/JNEUROSCI.2797-06.2006
|
| [15] |
Newell-Price J, Bertagna X, Grossman AB, et al. (2006) Cushing's syndrome. Lancet 367: 1605–1617. doi: 10.1016/S0140-6736(06)68699-6
|
| [16] |
Cobb J, O'Neill K, Milner J, et al. (2016) Density of GFAP-immunoreactive astrocytes is decreased in left hippocampi in major depressive disorder. Neuroscience 316: 209–220. doi: 10.1016/j.neuroscience.2015.12.044
|
| [17] | Vyas S, Rodrigues AJ, Silva JM, et al. (2016) Chronic stress and glucocorticoids: From neuronal plasticity to neurodegeneration. Neural Plast 2016: 6391686. |
| [18] |
Volterra A, Meldolesi J (2005) Astrocytes, from brain glue to communication elements: The revolution continues. Nat Rev Neurosci 6: 626–640. doi: 10.1038/nrn1722
|
| [19] | Koyama Y (2015) Functional alterations of astrocytes in mental disorders: Pharmacological significance as a drug target. Front Cell Neurosci 9: 261. |
| [20] | Carter BS (2013) Glucocorticoid regulation of the astrocyte transcriptome in vitro and in vivo. University of Michigan. |
| [21] |
Conti F, Debiasi S, Minelli A, et al. (1996) Expression of NR1 and NR2A/B subunits of the NMDA receptor in cortical astrocytes. Glia 17: 254–258. doi: 10.1002/(SICI)1098-1136(199607)17:3<254::AID-GLIA7>3.0.CO;2-0
|
| [22] |
Rajkowska G, Miguel-Hidalgo JJ, Wei J, et al. (1999) Morphometric evidence for neuronal and glial prefrontal cell pathology in major depression. Biol Psychiatry 45: 1085–1098. doi: 10.1016/S0006-3223(99)00041-4
|
| [23] |
Stockmeier CA, Mahajan GJ, Konick LC, et al. (2004) Cellular changes in the postmortem hippocampus in major depression. Biol Psychiatry 56: 640–650. doi: 10.1016/j.biopsych.2004.08.022
|
| [24] |
Rajkowska G, Stockmeier CA (2013) Astrocyte pathology in major depressive disorder: Insights from human postmortem brain tissue. Curr Drug Targets 14: 1225–1236. doi: 10.2174/13894501113149990156
|
| [25] |
Verkhratsky A, Rodriguez JJ, Parpura V (2013) Astroglia in neurological diseases. Future Neurol 8: 149–158. doi: 10.2217/fnl.12.90
|
| [26] |
Banasr M, Duman RS (2008) Glial Loss in the prefrontal cortex is sufficient to induce depressive-like behaviors. Biol Psychiatry 64: 863–870. doi: 10.1016/j.biopsych.2008.06.008
|
| [27] | Czeh B, Simon M, Schmelting B, et al. (2005) Astroglial plasticity in the hippocampus is affected by chronic psychosocial stress and concomitant fluoxetine treatment. Neuropsychopharmacology 31: 1616–1626. |
| [28] |
Banasr M, Chowdhury GMI, Terwilliger R, et al. (2010) Glial pathology in an animal model of depression: Reversal of stress-induced cellular, metabolic and behavioral deficits by the glutamate-modulating drug riluzole. Mol Psychiatry 15: 501–511. doi: 10.1038/mp.2008.106
|
| [29] |
Messing A, Brenner M (2003) GFAP: Functional implications gleaned from studies of genetically engineered mice. Glia 43: 87–90. doi: 10.1002/glia.10219
|
| [30] |
Miguel-Hidalgo JJ, Baucom C, Dilley G, et al. (2000) Glial fibrillary acidic protein immunoreactivity in the prefrontal cortex distinguishes younger from older adults in major depressive disorder. Biol Psychiatry 48: 861–873. doi: 10.1016/S0006-3223(00)00999-9
|
| [31] |
Bowley MP, Drevets WC, Ongur D, et al. (2002) Low glial numbers in the amygdala in major depressive disorder. Biol Psychiatry 52: 404–412. doi: 10.1016/S0006-3223(02)01404-X
|
| [32] | Polyakova M, Sander C, Arelin K, et al. (2015) First evidence for glial pathology in late life minor depression: S100B is increased in males with minor depression. Front Cell Neurosci 9: 406. |
| [33] |
Benton CS, Miller BH, Skwerer S, et al. (2012) Evaluating genetic markers and neurobiochemical analytes for fluoxetine response using a panel of mouse inbred strains. Psychopharmacology 221: 297–315. doi: 10.1007/s00213-011-2574-z
|
| [34] |
Chandley MJ, Szebeni K, Szebeni A, et al. (2013) Gene expression deficits in pontine locus coeruleus astrocytes in men with major depressive disorder. J Psychiatry Neurosci 38: 276–284. doi: 10.1503/jpn.120110
|
| [35] |
Bernard R, Kerman IA, Thompson RC, et al. (2011) Altered expression of glutamate signaling, growth factor, and glia genes in the locus coeruleus of patients with major depression. Mol Psychiatry 16: 634–646. doi: 10.1038/mp.2010.44
|
| [36] |
Seifert G, Schilling K, Steinhäuser C (2006) Astrocyte dysfunction in neurological disorders: A molecular perspective. Nat Rev Neurosci 7: 194. doi: 10.1038/nrn1870
|
| [37] |
Choudary P, Molnar M, Evans S, et al. (2005) Altered cortical glutamatergic and GABAergic signal transmission with glial involvement in depression. Proc Natl Acad Sci 102: 15653–15658. doi: 10.1073/pnas.0507901102
|
| [38] |
Sun JD, Liu Y, Yuan YH, et al. (2012) Gap junction dysfunction in the prefrontal cortex induces depressive-like behaviors in rats. Neuropsychopharmacology 37: 1305–1320. doi: 10.1038/npp.2011.319
|
| [39] |
Bechtholt-Gompf AJ, Walther HV, Adams MA, et al. (2010) Blockade of astrocytic glutamate uptake in rats induces signs of anhedonia and impaired spatial memory. Neuropsychopharmacology 35: 2049–2059. doi: 10.1038/npp.2010.74
|
| [40] |
de Vasconcellos-Bittencourt AP, Vendite DA, Nassif M, et al. (2011) Chronic stress and lithium treatments alter hippocampal glutamate uptake and release in the rat and potentiate necrotic cellular death after oxygen and glucose deprivation. Neurochem Res 36: 793–800. doi: 10.1007/s11064-011-0404-7
|
| [41] |
Almeida RF, Thomazi AP, Godinho GF, et al. (2010) Effects of depressive-like behavior of rats on brain glutamate uptake. Neurochem Res 35: 1164–1171. doi: 10.1007/s11064-010-0169-4
|
| [42] |
Bechtholt-Gompf AJ, Walther HV, Adams MA, et al. (2010) Blockade of astrocytic glutamate uptake in rats induces signs of anhedonia and impaired spatial memory. Neuropsychopharmacology 35: 2049–2059. doi: 10.1038/npp.2010.74
|
| [43] | Lapidus KA, Soleimani L, Murrough JW (2013) Novel glutamatergic drugs for the treatment of mood disorders. Neuropsychiatr Dis Treat 9: 1101–1112. |
| [44] |
Sanacora G, Treccani G, Popoli M (2012) Towards a glutamate hypothesis of depression: An emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacology 62: 63–77. doi: 10.1016/j.neuropharm.2011.07.036
|
| [45] |
Paslakis G, Gass P, Deuschle M (2011) [The role of the glutamatergic system in pathophysiology and pharmacotherapy for depression: Preclinical and clinical data]. Fortschr Neurol Psychiatr 79: 204–212. doi: 10.1055/s-0029-1245770
|
| [46] | Rial D, Lemos C, Pinheiro H, et al. (2015) Depression as a glial-based synaptic dysfunction. Front Cell Neurosci 9: 521. |
| [47] | Jo WK, Zhang Y, Emrich HM, et al. (2015) Glia in the cytokine-mediated onset of depression: fine tuning the immune response. Front Cell Neurosci 9: 268. |
| [48] | Brites D, Fernandes A (2015) Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front Cell Neurosci 9: 476. |
| [49] | Quesseveur G, Portal B, Basile JA, et al. (2015) Attenuated Levels of Hippocampal Connexin 43 and its Phosphorylation Correlate with Antidepressant- and Anxiolytic-Like Activities in Mice. Front Cell Neurosci 9: 490. |
| [50] | Jeanson T, Pondaven A, Ezan P, et al. (2015) Antidepressants Impact Connexin 43 Channel Functions in Astrocytes. Front Cell Neurosci 9: 495. |
| [51] |
Fuchs E, Flugge G (2003) Chronic social stress: Effects on limbic brain structures. Physiol Behav 79: 417–427. doi: 10.1016/S0031-9384(03)00161-6
|
| [52] |
Gould E, Tanapat P (1999) Stress and hippocampal neurogenesis. Biol Psychiatry 46: 1472–1479. doi: 10.1016/S0006-3223(99)00247-4
|
| [53] |
Rajkowska G, Miguel-Hidalgo JJ (2007) Gliogenesis and glial pathology in depression. CNS Neurol Disord Drug Targets 6: 219–233. doi: 10.2174/187152707780619326
|
| [54] |
Wang Q, Verweij EW, Krugers HJ, et al. (2014) Distribution of the glucocorticoid receptor in the human amygdala; changes in mood disorder patients. Brain Struct Funct 219: 1615–1626. doi: 10.1007/s00429-013-0589-4
|
| [55] |
Czeh B, Muller-Keuker JI, Rygula R, et al. (2007) Chronic social stress inhibits cell proliferation in the adult medial prefrontal cortex: Hemispheric asymmetry and reversal by fluoxetine treatment. Neuropsychopharmacology 32: 1490–1503. doi: 10.1038/sj.npp.1301275
|
| [56] |
Pekny M, Nilsson M (2005) Astrocyte activation and reactive gliosis. Glia 50: 427–434. doi: 10.1002/glia.20207
|
| [57] |
Rozovsky I, Laping NJ, Krohn K, et al. (1995) Transcriptional regulation of glial fibrillary acidic protein by corticosterone in rat astrocytes in vitro is influenced by the duration of time in culture and by astrocyte-neuron interactions. Endocrinology 136: 2066–2073. doi: 10.1210/endo.136.5.7720656
|
| [58] |
O'Callaghan JP, Brinton RE, Mcewen BS (1991) Glucocorticoids regulate the synthesis of glial fibrillary acidic protein in intact and adrenalectomized rats but do not affect its expression following brain injury. J Neurochem 57: 860–869. doi: 10.1111/j.1471-4159.1991.tb08230.x
|
| [59] | Bridges N, Slais K, Sykova E (2008) The effects of chronic corticosterone on hippocampal astrocyte numbers: A comparison of male and female Wistar rats. Acta Neurobiol Exp 68: 131–138. |
| [60] |
Lambert KG, Gerecke KM, Quadros PS, et al. (2000) Activity-stress increases density of GFAP-immunoreactive astrocytes in the rat hippocampus. Stress 3: 275–284. doi: 10.3109/10253890009001133
|
| [61] |
Leventopoulos M, Ruedi-Bettschen D, Knuesel I, et al. (2007) Long-term effects of early life deprivation on brain glia in Fischer rats. Brain Res 1142: 119–126. doi: 10.1016/j.brainres.2007.01.039
|
| [62] |
Fields RD, Stevens-Graham B (2002) New insights into neuron-glia communication. Science 298: 556–562. doi: 10.1126/science.298.5593.556
|
| [63] |
Vardimon L, Ben-Dror I, Avisar N, et al. (1999) Glucocorticoid control of glial gene expression. J Neurobiol 40: 513–527. doi: 10.1002/(SICI)1097-4695(19990915)40:4<513::AID-NEU8>3.0.CO;2-D
|
| [64] |
Conrad CD (2006) What is the functional significance of chronic stress-induced CA3 dendritic retraction within the hippocampus? Behav Cogn Neurosci Rev 5: 41–60. doi: 10.1177/1534582306289043
|
| [65] |
Autry AE, Grillo CA, Piroli GG, et al. (2006) Glucocorticoid regulation of GLT-1 glutamate transporter isoform expression in the rat hippocampus. Neuroendocrinology 83: 371–379. doi: 10.1159/000096092
|
| [66] |
Horner HC, Packan DR, Sapolsky RM (1990) Glucocorticoids inhibit glucose transport in cultured hippocampal neurons and glia. Neuroendocrinology 52: 57–64. doi: 10.1159/000125539
|
| [67] |
Ritchie LJ, De Butte M, Pappas BA (2004) Chronic mild stress exacerbates the effects of permanent bilateral common carotid artery occlusion on CA1 neurons. Brain Res 1014: 228–235. doi: 10.1016/j.brainres.2004.04.036
|
| [68] |
Kroll S, El-Gindi J, Thanabalasundaram G, et al. (2009) Control of the blood-brain barrier by glucocorticoids and the cells of the neurovascular unit. Ann N Y Acad Sci 1165: 228–239. doi: 10.1111/j.1749-6632.2009.04040.x
|
| [69] |
Donato R (2001) S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int J Biochem Cell Biol 33: 637–668. doi: 10.1016/S1357-2725(01)00046-2
|
| [70] |
Bruschettini M, van den Hove DL, Gazzolo D, et al. (2005) A single course of antenatal betamethasone reduces neurotrophic factor S100B concentration in the hippocampus and serum in the neonatal rat. Brain Res Dev Brain Res 159: 113–118. doi: 10.1016/j.devbrainres.2005.07.003
|
| [71] |
Margis R, Zanatto VC, Tramontina F, et al. (2004) Changes in S100B cerebrospinal fluid levels of rats subjected to predator stress. Brain Res 1028: 213–218. doi: 10.1016/j.brainres.2004.09.013
|
| [72] |
Scaccianoce S, Del BP, Pannitteri G, et al. (2004) Relationship between stress and circulating levels of S100B protein. Brain Res 1004: 208–211. doi: 10.1016/j.brainres.2004.01.028
|
| [73] |
Niu H, Hinkle DA, Wise PM (1997) Dexamethasone regulates basic fibroblast growth factor, nerve growth factor and S100beta expression in cultured hippocampal astrocytes. Brain Res Mol Brain Res 51: 97–105. doi: 10.1016/S0169-328X(97)00221-0
|
| [74] |
Bazargani N, Attwell D (2016) Astrocyte calcium signaling: the third wave. Nat Neurosci 19: 182–189. doi: 10.1038/nn.4201
|
| [75] |
Simard M, Couldwell WT, Zhang W, et al. (1999) Glucocorticoids-potent modulators of astrocytic calcium signaling. Glia 28: 1–12. doi: 10.1002/(SICI)1098-1136(199910)28:1<1::AID-GLIA1>3.0.CO;2-4
|
| [76] |
Mcleod JD, Bolton C (1995) Dexamethasone induces an increase in intracellular and membrane-associated lipocortin-1 (annexin-1) in rat astrocyte primary cultures. Cell Mol Neurobiol 15: 193–205. doi: 10.1007/BF02073328
|
| [77] |
Lindholm D, Castren E, Hengerer B, et al. (1992) Differential Regulation of Nerve Growth Factor (NGF) Synthesis in Neurons and Astrocytes by Glucocorticoid Hormones. Eur J Neurosci 4: 404–410. doi: 10.1111/j.1460-9568.1992.tb00889.x
|
| [78] |
Yuan SY, Liu J, Zhou J, et al. (2016) AMPK Mediates Glucocorticoids Stress-Induced Downregulation of the Glucocorticoid Receptor in Cultured Rat Prefrontal Cortical Astrocytes. PLoS One 11: e0159513. doi: 10.1371/journal.pone.0159513
|
| [79] |
Chen J, Wang ZZ, Zuo W, et al. (2016) Effects of chronic mild stress on behavioral and neurobiological parameters-Role of glucocorticoid. Horm Behav 78: 150–159. doi: 10.1016/j.yhbeh.2015.11.006
|
| [80] |
Karisetty BC, Joshi PC, Kumar A, et al. (2017) Sex differences in the effect of chronic mild stress on mouse prefrontal cortical BDNF levels: A role of major ovarian hormones. Neuroscience 356: 89–101. doi: 10.1016/j.neuroscience.2017.05.020
|
| [81] |
Bender CL, Calfa GD, Molina VA (2016) Astrocyte plasticity induced by emotional stress: A new partner in psychiatric physiopathology? Prog Neuropsychopharmacol Biol Psychiatry 65: 68–77. doi: 10.1016/j.pnpbp.2015.08.005
|
| [82] |
Anacker C, Cattaneo A, Luoni A, et al. (2013) Glucocorticoid-related molecular signaling pathways regulating hippocampal neurogenesis. Neuropsychopharmacology 38: 872–883. doi: 10.1038/npp.2012.253
|
| [83] |
Van DL, Steinbusch HW, Bruschettini M, et al. (2006) Prenatal stress reduces S100B in the neonatal rat hippocampus. Neuroreport 17: 1077–1080. doi: 10.1097/01.wnr.0000223391.74575.c9
|
| [84] |
Nichols NR, Agolley D, Zieba M, et al. (2005) Glucocorticoid regulation of glial responses during hippocampal neurodegeneration and regeneration. Brain Res Rev 48: 287–301. doi: 10.1016/j.brainresrev.2004.12.019
|
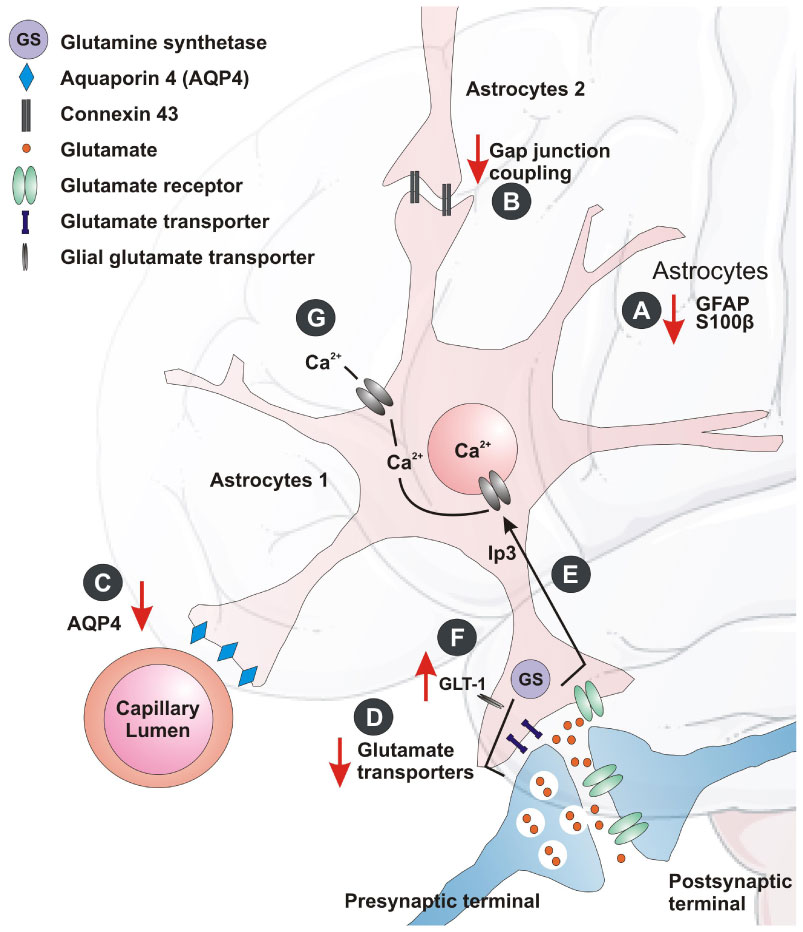
Figures(1) / Tables(1)

Pranav Chintamani Joshi, Sugato Benerjee. Effects of glucocorticoids in depression: Role of astrocytes[J]. AIMS Neuroscience, 2018, 5(3): 200-210. doi: 10.3934/Neuroscience.2018.3.200







 DownLoad:
DownLoad: