Citation: Juliet Lee. Insights into cell motility provided by the iterative use of mathematical modeling and experimentation[J]. AIMS Biophysics, 2018, 5(2): 97-124. doi: 10.3934/biophy.2018.2.97

| [1] |
Pollard TD, Borisy GG (2003) Cellular motility driven by assembly and disassembly of actin filaments. Cell 112: 453–465. doi: 10.1016/S0092-8674(03)00120-X

|
| [2] |
Pollard TD (2003) The cytoskeleton, cellular motility and the reductionist agenda. Nature 422: 741–745. doi: 10.1038/nature01598
|
| [3] |
Holmes WR, Edelsteinkeshet L (2012) A comparion of computational models for eukaryotic cell hape and motility. PLoS Comput Biol 8: e1002793. doi: 10.1371/journal.pcbi.1002793
|
| [4] |
Danuser G, Allard J, Mogliner A (2013) Mathematical modeling of eukaryotic cell migration: Insights beyond experiments. Annu Rev Cell Dev Biol 29: 501–528. doi: 10.1146/annurev-cellbio-101512-122308
|
| [5] |
Mogiler A (2009) Mathematics of cell motility: Have we got its number? J Math Biol 58: 105–134. doi: 10.1007/s00285-008-0182-2
|
| [6] | Mogilner A, Keren AK (2009) The shape of motile cells. Curr Biol 15: 762–771. |
| [7] | Oelz D, Schmeiser C (2011) Simulation of lamellipodial fragments. J Math Biol 64: 513–528. |
| [8] | Adler Y, Givli S (2013) Closing the loop: Lamellipodia dynamics from the perspective of front propagation. Phys Rev E 88: 042708. |
| [9] |
Recho P, Putelat T, Truskinovsky L (2013) Contraction-driven cell motility. Phys Rev Lett 111: 108102. doi: 10.1103/PhysRevLett.111.108102
|
| [10] |
Tjhung E, Tiribocchi A, Marenduzzo D, et al. (2015) A minimal physical model captures the shapes of crawling cells. Nat Commun 6: 5420. doi: 10.1038/ncomms6420
|
| [11] | Ambrosi D, Zanzottera A (2016) Mechanics and polarity in cell motility. Physica D 330: 58–66. |
| [12] |
Raynaud F, Ambühl ME, Gabella C, et al. (2016) Minimal models for spontaneous cell polarization and edge activity in oscillating, rotating and migrating cells. Nat Phys 12: 367–374. doi: 10.1038/nphys3615
|
| [13] |
Pollard TD, Cooper JA (2009) Actin, a central player in cell shape and movement. Science 326: 1208–1212. doi: 10.1126/science.1175862
|
| [14] |
Rafelski SM, Theriot JA (2004) Crawling toward a unified model of cell motility: Spatial and temporal regulation of actin dynamics. Annu Rev Biochem 73: 209–239. doi: 10.1146/annurev.biochem.73.011303.073844
|
| [15] | Mogilner A (2006) On the edge: Modeling protrusion. Curr Opin Cell Biol 18: 32–39. |
| [16] |
Svitkina TM, Verkhovsky AB, Mcquade KM, et al. (1997) Analysis of the actin-myosin II system in fish epidermal keratocytes: Mechanism of cell body translocation. J Cell Biol 139: 397–415. doi: 10.1083/jcb.139.2.397
|
| [17] |
Clainche CL, Carlier MF (2008) Regulation of actin assembly associated with protrusion and adhesion in cell migration. Physiol Rev 88: 489–513. doi: 10.1152/physrev.00021.2007
|
| [18] |
Vincente-Manzaneres M, Choi CK, Horwitz AR (2009) Integrins in cell migration-the actin connection. J Cell Sci 122: 199–206. doi: 10.1242/jcs.018564
|
| [19] |
Gardel ML, Schneider IC, Aratyn-Schaus Y, et al. (2010) Mechanical integration of actin and adhesion dynamics in cell migration. Annu Rev Cell Dev Biol 26: 315–333. doi: 10.1146/annurev.cellbio.011209.122036
|
| [20] |
Suter DM, Forscher P (2000) Substrate-cytoskeletal coupling as a mechanism for the regulation of growth cone motility and guidance. Dev Neurobiol 44: 97–113. doi: 10.1002/1097-4695(200008)44:2<97::AID-NEU2>3.0.CO;2-U
|
| [21] | Vincente-Manzaneres M, Ma X, Adelstein RS, et al. (2009) Non-muscle myosin II takes centre stage in cell adhesion and migration. Nat Rev Mol Cell Biol 10: 788–790. |
| [22] |
Kirfel G, Rigort A, Borm B, et al. (2004) Cell migration: Mechanisms of rear detachment and the formation of migration tracks. Eur J Cell Biol 83: 717–724. doi: 10.1078/0171-9335-00421
|
| [23] |
Lee J, Ishihara A, Oxford G, et al. (1999) Regulation of cell movement is mediated by stretch-activated calcium channels. Nature 400: 382–386. doi: 10.1038/22578
|
| [24] |
Huttenlocher A, Palecek SP, Lu Q, et al. (1997) Regulation of cell migration by the calcium-dependent protease calpain. J Biol Chem 272: 32719–32722. doi: 10.1074/jbc.272.52.32719
|
| [25] |
Wolfenson H, Bershadsky A, Henis Y, et al. (2011) Actomyosin-generated tension controls the molecular kinetics of focal adhesions. J Cell Sci 124: 1425–1432. doi: 10.1242/jcs.077388
|
| [26] | Verkhovsky AB, Svitkina T, Borisy GG (1999) Self-polarization and directional motility of cytoplasm. Curr Biol 9: 11–20. |
| [27] |
Ridley AJ, Horwitz AR (2003) Cell migration: Integrating signals from front to back. Science 302: 1704–1709. doi: 10.1126/science.1092053
|
| [28] | Friedl P, Sahai E, Weiss S, et al. (2012) New dimensions in cell migration. Nat Rev Mol Cell Biol 13: 743–747. |
| [29] |
Lo CM, Wang HB, Dembo M, et al. (2000) Cell movement is guided by the rigidity of the substrate. Biophys J 79: 144–152. doi: 10.1016/S0006-3495(00)76279-5
|
| [30] |
Abercrombie M (1961) The bases of locomotory behaviour of fibroblasts. Exp Cell Res 8: 188. doi: 10.1016/0014-4827(61)90348-2
|
| [31] |
Allan RB, Wilkinson PC (1978) A visual analysis of chemotactic and chemokinetic locomotion of human neutrophil leukocytes. Exp Cell Res 111: 191–203. doi: 10.1016/0014-4827(78)90249-5
|
| [32] | Lee J, Ishihara A, Jacobson K (1993) The fish epidermal keratocyte as a model system for the study of cell locomotion. Symp Soc Exp Biol 47: 73–89. |
| [33] |
Tranquillo RT, Lauffenburger DA, Zigmond SH (1988) A stochastic model for leukocyte random motility and chemotaxis bases on receptor binding fluctuations. J Cell Biol 106: 303–309. doi: 10.1083/jcb.106.2.303
|
| [34] | Mogilner A, Verzi DW (2003) A Simple 1-D physical model for the crawling nematode sperm cell. J Stat Phys 110: 1169–1189. |
| [35] |
Herant M, Dembo M (2010) Form and function in cell motility: From fibroblasts to keratocytes. Biophys J 98: 1408–1417. doi: 10.1016/j.bpj.2009.12.4303
|
| [36] |
Satulovsky J, Lui R, Wang Yl (2008) Exploring the control circuit of cell migration by mathematical modeling. Biophys J 94: 3671–3683. doi: 10.1529/biophysj.107.117002
|
| [37] |
Grimm HP, Verkhovsky AB, Mogilner A, et al. (2003) Analysis of actin dynamics at the leading edge of crawling cells: Implications for the shape of the keratocyte lamellipodia. Eur Biophys J 32: 563–577. doi: 10.1007/s00249-003-0300-4
|
| [38] | Hellewell SB, Taylor DL (1979) The contractile basis of amoeboid movements. VI. The solation-contraction coupling hypothesis. J Cell Biol 83: 633–648. |
| [39] |
Chen WT (1979) Induction of spreading during fibroblast movement. J Cell Biol 81: 684–691. doi: 10.1083/jcb.81.3.684
|
| [40] |
Harris A, Dunn G (1972) Centripetal transport of attached particles on both surfaces of moving fibroblasts. Exp Cell Res 73: 519–523. doi: 10.1016/0014-4827(72)90084-5
|
| [41] |
Lee J, Ishihara A, Teriot JA, et al. (1993) Principles of locomotion for simple-shaped cells. Nature 362: 167–171. doi: 10.1038/362167a0
|
| [42] |
Keren K, Pincus Z, Allen GM, et al. (2008) Mechanism of shape determination in motile cells. Nature 453: 475–480. doi: 10.1038/nature06952
|
| [43] |
Rubenstein B, Jacobson K, Moglilner A (2005) Multiscale two-dimensional modeling of a motile simple-shaped cell. Multiscale Model Sim 3: 413–439. doi: 10.1137/04060370X
|
| [44] | Barnhart EL, Lee KC, Keren K, et al. (2011) An adhesion-dependent switch between mechanisms that determine motile cell shape. PLoS Biol 9: 1–16. |
| [45] |
Wolgemuth C, Stajic J, Mogliner A (2011) Redundant mechanisms for stable cell locomotion revealed by minimal models. Biophys J 101: 545–553. doi: 10.1016/j.bpj.2011.06.032
|
| [46] | Bottino D, Moglilner A, Roberts T, et al. (2001) How nematode sperm crawl. J Cell Sci 115: 367–384. |
| [47] |
Roberts T, Stewart M (2000) Acting like actin: The dynamics of the nematode major sperm protein (MSP) cytoskeleton indicate a push-pull mechanism for amoeboid cell motility. J Cell Biol 149: 7–12. doi: 10.1083/jcb.149.1.7
|
| [48] |
Janson LW, Kolega J, Taylor DL (1991) Modulation of contraction by gelation/solation in a reconstituted motile model. J Cell Biol 114: 1005–1015. doi: 10.1083/jcb.114.5.1005
|
| [49] |
Forscher P, Smith SJ (1988) Actions of cytochalasins on the organization of actin filaments and microtubules in a neuronal growth cone. J Cell Biol 107: 1505–1516. doi: 10.1083/jcb.107.4.1505
|
| [50] |
Theriot JA, Mitchison TJ (1991) Actin microfilament dynamics in locomoting cells. Nature 352: 126–131. doi: 10.1038/352126a0
|
| [51] |
Harris AK, Wild P, Stopal D (1980) Silicone rubber substrata: A new wrinkle in the study of cell locomotion. Science 208: 177–179. doi: 10.1126/science.6987736
|
| [52] |
Wang JHC, Shang JS (2007) Cell traction force and measurement methods. Biomech Model Mechan 6: 361–371. doi: 10.1007/s10237-006-0068-4
|
| [53] |
Beningo KA, Dembo M, Kverina I, et al. (2001) Nascent focal adhesions are responsible for the generation of strong propulsive forces in migrating fibroblasts. J Cell Biol 153: 881–887. doi: 10.1083/jcb.153.4.881
|
| [54] |
Hind LE, Dembo M, Hammer DA (2015) Macrophage motility is driven by frontal-towing with a force magnitude dependent on substrate stiffness. Integr Biol 7: 447–453. doi: 10.1039/C4IB00260A
|
| [55] |
Doyle A, Marganski W, Lee J (2004) Calcium transients induce spatially coordinated increases in traction force during the movement of fish keratocytes. J Cell Sci 117: 2203–2214. doi: 10.1242/jcs.01087
|
| [56] |
Loisel TP, Boujemaa R, Pantaloni D, et al. (1999) Reconstitution of actin-based motility of Listeria and Shigella using pure proteins. Nature 401: 613–616. doi: 10.1038/44183
|
| [57] |
Roy P, Rajfur Z, Jones D (2001) Local photorelease of caged thymosin beta 4 in locomoting keratocytes causes cell turning. J Cell Biol 153: 1035–1047. doi: 10.1083/jcb.153.5.1035
|
| [58] |
DiMilla PA, Barbee K, Lauffenburger DA (1991) Mathematical model for the effects of adhesion and mechanics on cell migration speed. Biophys J 60: 15–37. doi: 10.1016/S0006-3495(91)82027-6
|
| [59] |
Gupton SL, Waterman-Storer CM (2006) Spatiotemporal feedback between actomyosin and focal-adhesion systems optimizes rapid cell migration. Cell 125: 1361–1374. doi: 10.1016/j.cell.2006.05.029
|
| [60] | Jurado C, Haserick JR, Lee J (2005) Slipping or gripping? Fluorescent speckle microscopy in fish keratocytes reveals two different mechanisms for generating a retrograde flow of actin. Mol Biol Cell 16: 507–518. |
| [61] |
Wilson CA, Tsuchida MA, Allen GM, et al. (2010) Myosin II contributes to cell-scale actin network treadmilling through network disassembly. Nature 465: 373–377. doi: 10.1038/nature08994
|
| [62] |
Fournier MF, Sauser S, Ambrosi D, et al. (2010) Force transimission in migrating cells. J Cell Biol 188: 287–297. doi: 10.1083/jcb.200906139
|
| [63] |
Lee J, Leonard M, Oliver T, et al. (1994) Traction forces generated by locomoting keratocytes. J Cell Biol 127: 1957–1964. doi: 10.1083/jcb.127.6.1957
|
| [64] |
Hartwell LH, Hopfield JJ, Leibler S, et al. (1999) From molecular to modular cell biology. Nature 402: C47–C52. doi: 10.1038/35011540
|
| [65] |
Italiano JE, Stewart M, Roberts TM (2001) How the assembly dynamics of the nematode major sperm protein generate amoeboid cell motility. Int Rev Cytol 202: 1–34. doi: 10.1016/S0074-7696(01)02002-2
|
| [66] |
Friedl P, Wolf K (2010) Plasticity of cell migration: A multiscale tuning model. J Cell Biol 188: 11–19. doi: 10.1083/jcb.200909003
|
| [67] |
Doyle AD, Lee J (2005) Cyclic changes in keratocyte speed and traction stress arise from Ca2+-dependent regulation of cell adhesiveness. J Cell Sci 118: 369–379. doi: 10.1242/jcs.01590
|
| [68] |
Paku S, Tóvári J, Lörincz Z, et al. (2003) Adhesion dynamics and cytoskeletal structure of gliding human fibrosarcoma cells: A hypothetical model of cell migration. Exp Cell Res 290: 246–253. doi: 10.1016/S0014-4827(03)00334-3
|
| [69] |
Huang C, Rajfur Z, Borchers C, et al. (2003) JNK phosphorylates paxillin and regulates cell migration. Nature 424: 219–223. doi: 10.1038/nature01745
|
| [70] |
Asano Y, Mizuno T, Kon T, et al. (2004) Keratocyte-like locomotion in amiB-null Dictyostelium cells. Cell Motil Cytoskel 59: 17–27. doi: 10.1002/cm.20015
|
| [71] |
Ma L, Janetopoulos C, Yang L, et al. (2004) Two complementary, local excitation, global inhibition mechanisms acting in parallel can explain the chemoattractant-induced regulation of PI(3,4,5)P-3 response in Dictyostelium cells. Biophys J 87: 3764–3774. doi: 10.1529/biophysj.104.045484
|
| [72] |
Yam PT, Wilson CA, Ji L, et al. (2007) Actin-myosin network reorganization breaks symmetry at the cell rear to spontaneously initiate polarized cell motility. J Cell Biol 178: 1207–1221. doi: 10.1083/jcb.200706012
|
| [73] | Lee J, Jacobson K (1997) The composition and dynamics of cell-substratum adhesions in locomoting fish keratocytes. J Cell Sci 110: 2833–2844. |
| [74] |
Anderson KI, Cross R (2000) Contact dynamics during keratocyte motility. Curr Biol 10: 253–260. doi: 10.1016/S0960-9822(00)00357-2
|
| [75] |
Morin TR, Ghassemzadeh SA, Lee J (2014) Traction force microscopy in rapidly moving cells reveals separate roles for ROCK and MLCK in the mechanics of retraction. Exp Cell Res 326: 280–294. doi: 10.1016/j.yexcr.2014.04.015
|
| [76] |
Shao D, Levine H, Rappel WJ (2012) Coupling actin flow, adhesion, and morphology in a computational cell motiliy model. PNAS 109: 6851–6856. doi: 10.1073/pnas.1203252109
|
| [77] |
Ziebert F, Aranson IS (2014) Modular approach for modeling cell motility. Eur Phys J-Spec Top 223: 1265–1277. doi: 10.1140/epjst/e2014-02190-2
|
| [78] |
Camely BA, Zhao Y, Li B, et al. (2017) Crawling and turning in a minimal reaction-diffusion cell motility model: Coupling cell shape and biochemistry. Phys Rev E 95: 012401. doi: 10.1103/PhysRevE.95.012401
|
| [79] |
Barnhart EL, Allen GM, Julicher F, et al. (2010) Bipedal locomotion in crawling cells. Biophys J 98: 933–942. doi: 10.1016/j.bpj.2009.10.058
|
| [80] |
Sanz-Morena V, Gadea G, Paterson H, et al. (2008) Rac activation and inactivation control plasticity of tumor cell movement. Cell 135: 510–523. doi: 10.1016/j.cell.2008.09.043
|
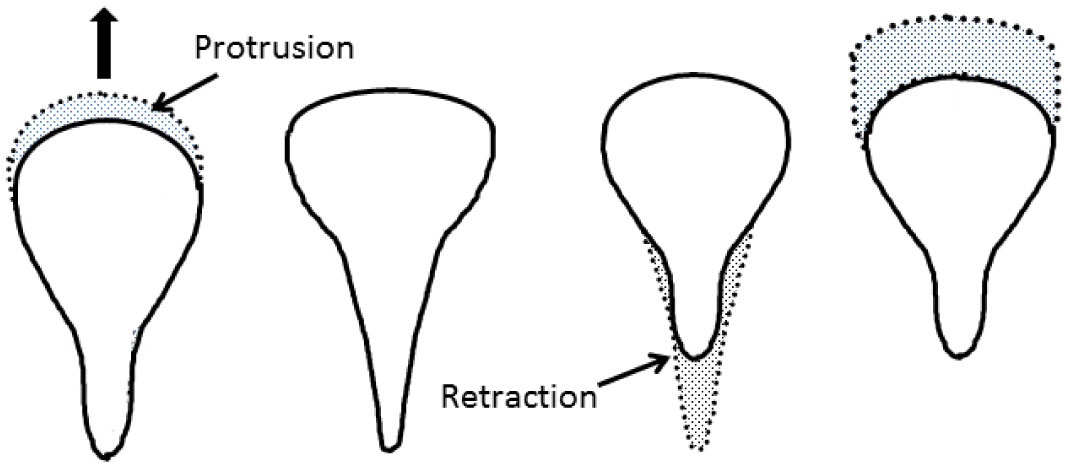
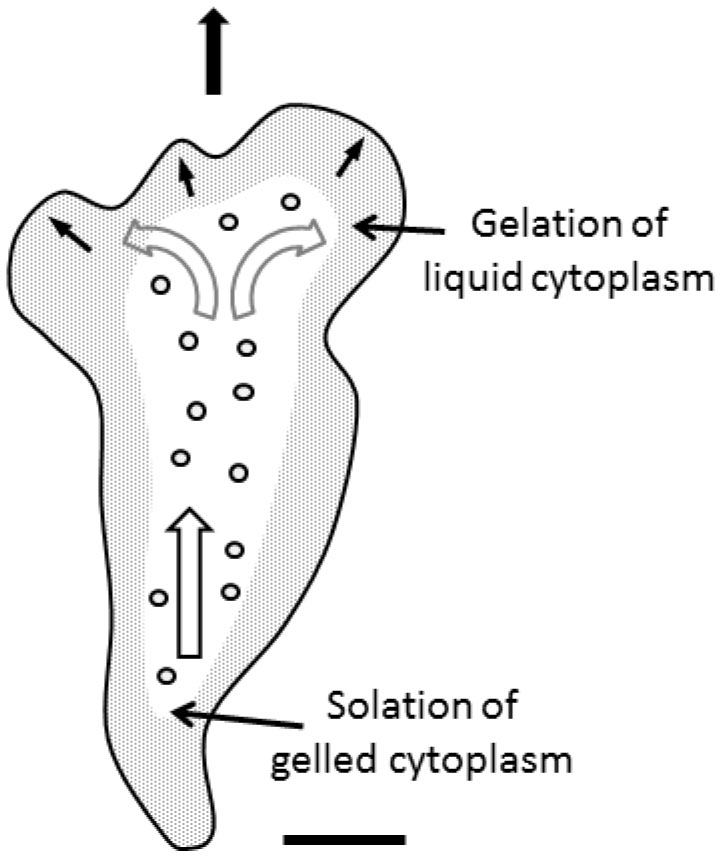
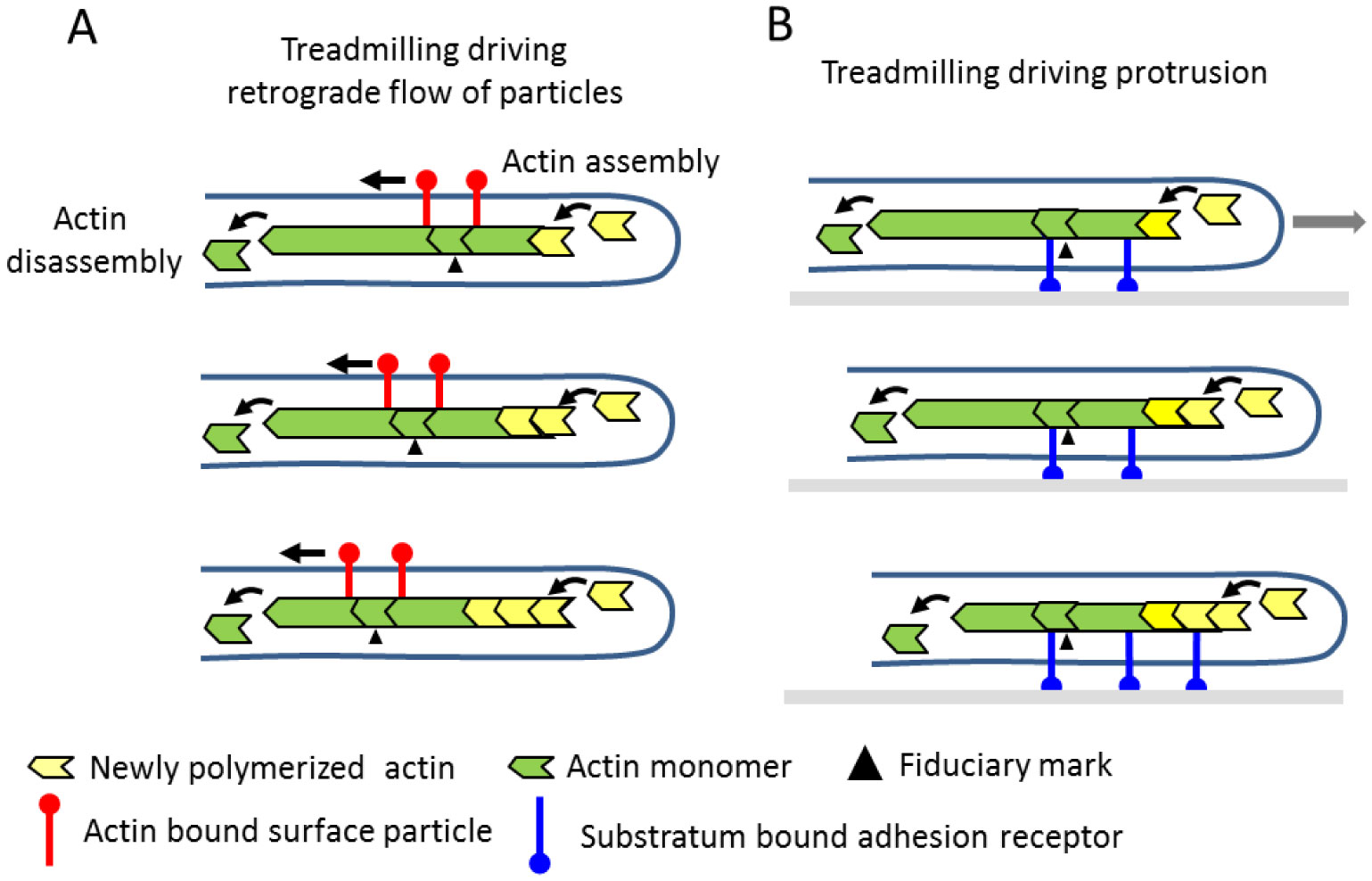
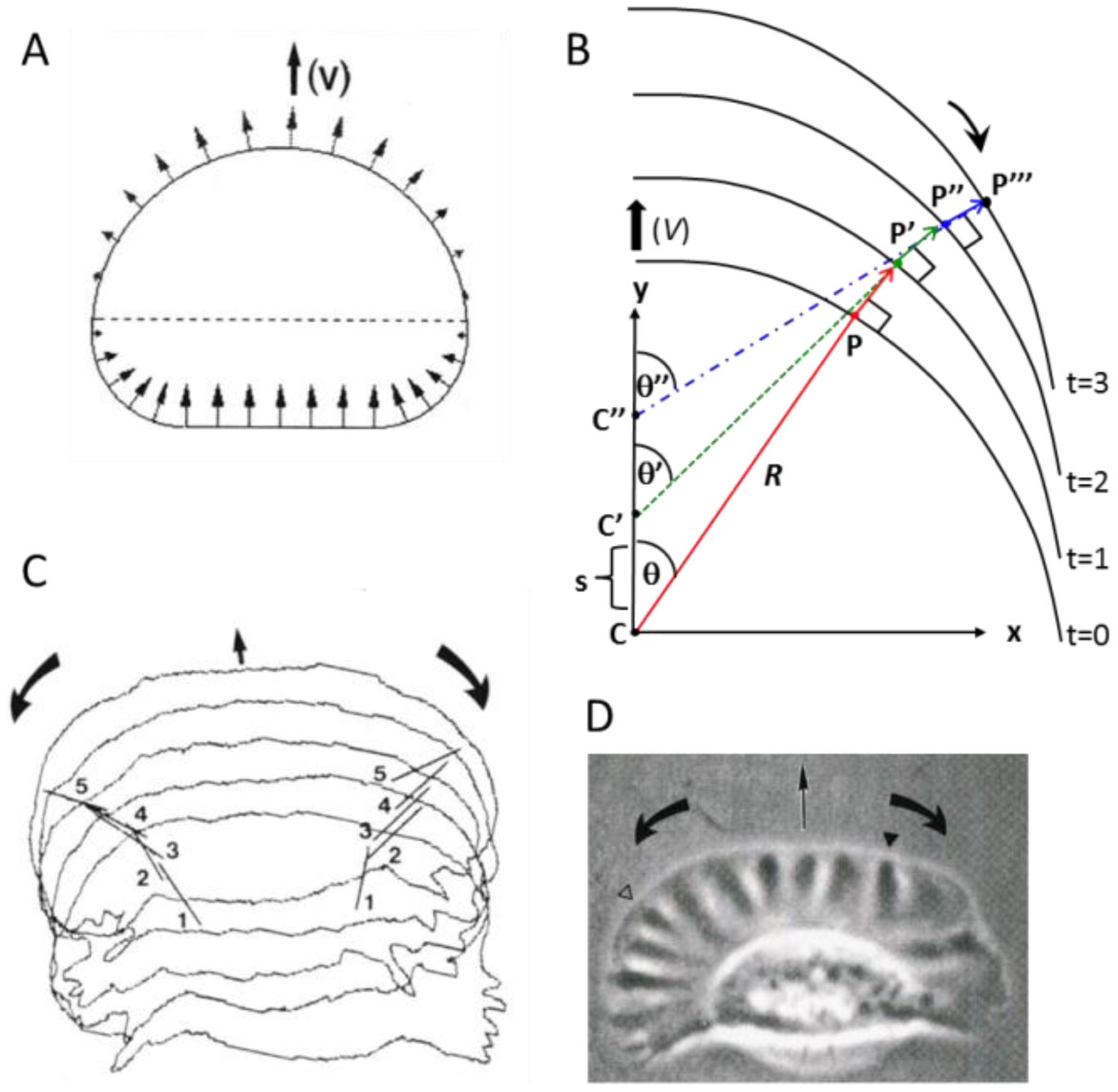
Figures(9) / Tables(1)

Juliet Lee. Insights into cell motility provided by the iterative use of mathematical modeling and experimentation[J]. AIMS Biophysics, 2018, 5(2): 97-124. doi: 10.3934/biophy.2018.2.97







 DownLoad:
DownLoad: