Citation: Malin Bomberg, Mari Raulio, Sirpa Jylhä, Carsten W. Mueller, Carmen Höschen, Pauliina Rajala, Lotta Purkamo, Riikka Kietäväinen, Lasse Ahonen, Merja Itävaara. CO2 and carbonate as substrate for the activation of the microbial community in 180 m deep bedrock fracture fluid of Outokumpu Deep Drill Hole, Finland[J]. AIMS Microbiology, 2017, 3(4): 846-871. doi: 10.3934/microbiol.2017.4.846

| [1] | Purkamo L, Bomberg M, Nyyssönen M, et al. (2015) Heterotrophic communities supplied by ancient organic carbon predominate in deep Fennoscandian bedrock fluids. Microb Ecol: 319–332. |
| [2] | Kukkonen IT (2011) Introduction to the Outokumpu Deep Drilling Project 2003–2010, In: Kukkonen IT, Editor, Outokumpu Deep Drilling Project 2003–2010, Special Paper 51, Espoo: Geological Survey of Finland, 11–16. |
| [3] |
Kietäväinen R, Ahonen L, Kukkonen IT, et al. (2013) Characterisation and isotopic evolution of saline waters of the Outokumpu Deep Drill Hole, Finland-Implications for water origin and deep terrestrial biosphere. Appl Geochem 32: 37–51. doi: 10.1016/j.apgeochem.2012.10.013

|
| [4] | Kietäväinen R, Ahonen L, Niinikoski P, et al. (2017) Abiotic and biotic controls on methane formation down to 2.5 km depth within the Precambrian Fennoscandian Shield. Geochim Cosmochim Acta 202: 124–145. |
| [5] |
Nyyssönen M, Hultman J, Ahonen L, et al. (2014) Taxonomically and functionally diverse microbial communities in deep crystalline rocks of the Fennoscandian shield. ISME J 8: 126–138. doi: 10.1038/ismej.2013.125
|
| [6] | Rajala P, Bomberg M, Kietäväinen R, et al. (2015) Deep subsurface microbes rapidly reactivate in the presence of C-1 Compounds. Microorganisms 3: 17–33. |
| [7] | Rajala P, Bomberg M (2017) Reactivation of deep subsurface microbial community in response to methane or methanol amendment. Front Microbiol: 8. |
| [8] | Purkamo L, Bomberg M, Nyyssönen M, et al. (2013) Retrieval and analysis of authentic microbial communities from packer-isolated deep crystalline bedrock fractures: evaluation of the method and time of sampling. FEMS Microbiol Ecol 85: 324–337. |
| [9] | Itävaara M, Nyyssönen M, Bomberg M, et al. (2011) Microbiological sampling and analysis of Outokumpu Deep Borehole biosphere in 2007–2008, In: Kukkonen IT, Editor, Outokumpu Deep Drilling Project 2003–2010, Special Paper 51, Espoo: Geological Survey of Finland, 199–206. |
| [10] |
Itävaara M, Nyyssönen M, Kapanen A, et al. (2011) Characterization of bacterial diversity down to a depth of 1500 m of the Outokumpu deep borehole. FEMS Microbiol Ecol 77: 295–309. doi: 10.1111/j.1574-6941.2011.01111.x
|
| [11] | Ahonen L, Kietäväinen R, Kortelainen N, et al. (2011) Hydrogeological characteristics of the Outokumpu Deep Drill Hole, In: Kukkonen IT, Editor, Outokumpu Deep Drilling Project 2003–2010, Special Paper 51, Espoo: Geological Survey of Finland, 151–168. |
| [12] |
Weisburg WG, Barns SM, Pelletier DA, et al. (1991) 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol 173: 697–703. doi: 10.1128/jb.173.2.697-703.1991
|
| [13] |
Nyyssönen M, Bomberg M, Kapanen A, et al. (2012) Methanogenic and sulphate-reducing microbial communities in deep groundwater of crystalline rock fractures in Olkiluoto, Finland. Geomicrobiol J 29: 863–878. doi: 10.1080/01490451.2011.635759
|
| [14] | Herlemann DP, Labrenz M, Jürgens K, et al. (2011) Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J 5: 1571–1579. |
| [15] |
Klindworth A, Pruesse E, Schweer T, et al. (2013) Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res 41: 1–11. doi: 10.1093/nar/gks1039
|
| [16] | Buee M, Reich M, Murat C, et al. (2009) 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol 184: 449–456. |
| [17] | Quast C, Pruesse E, Yilmaz P, et al. (2013) The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res 41: D590–D596. |
| [18] |
Kõljalg U, Nilsson RH, Abarenkov K, et al. (2013) Towards a unified paradigm for sequence-based identification of Fungi. Mol Ecol 22: 5271–5277. doi: 10.1111/mec.12481
|
| [19] |
McMurdie P, Holmes S (2015) Shiny-phyloseq: web application for interactive microbiome analysis with provenance tracking. Bioinformatics 31: 282–283. doi: 10.1093/bioinformatics/btu616
|
| [20] | R Development Core Team, R: A Language and Environment for Statistical Computing, 2013. Available from: http://www.r-project.org/. |
| [21] | Aragno M (1998) The aerobic hydrogen-oxidizing (Knallgas) bacteria, In: Burlage RS, Atlas R, Stahl D, et al., Editors, Techniques in microbial ecology, 1 Eds., New York: Oxford University Press, 79. |
| [22] | Nübel U, Engelen B, Felske A, et al. (1996) Sequence heterogeneities of genes encoding 16S rRNAs in Paenibacillus polymyxa detected by temperature gradient gel electrophoresis. J Bacteriol 78: 5636–5643. |
| [23] |
Guidon S, Gascuel O (2003) A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst Biol 52: 696–704. doi: 10.1080/10635150390235520
|
| [24] | Jukes TH, Cantor CR (1969) Evolution of protein molecules, In: Munro HN, Editor, Mammalian protein metabolism, 1 Eds., New York: Academic Press, 21–132. |
| [25] |
D'Hondt S, Rutherford S, Spivack JS (2002) Metabolic activity of subsurface life in deep-sea sediments. Science 295: 2067–2070. doi: 10.1126/science.1064878
|
| [26] |
Parkes RJ, Cragg BA, Banning N, et al. (2007) Biogeochemistry and biodiversity of methane cycling in subsurface marine sediments (Skagerrak, Denmark). Environ Microbiol 9: 1146–1161. doi: 10.1111/j.1462-2920.2006.01237.x
|
| [27] |
Berman T, Kaplan B, Chava S, et al. (2001) Metabolically active bacteria in Lake Kinneret. Aquat Microb Ecol 23: 213–224. doi: 10.3354/ame023213
|
| [28] | Senjarini K, Karsten U, Schumann R (2013) Application of fluorescence markers for the diagnosis of bacterial abundance and viability in aquatic ecosystem. J Microbiol Res 3: 143–147. |
| [29] |
Proctor LM, Souza AC (2001) Method for enumeration of 5-cyano-2,3-ditoyl tetrazolium chloride (CTC)-active cells and cell-specific CTC activity of benthic bacteria in riverine, estuarine and coastal sediments. J Microbiol Meth 43: 213–222. doi: 10.1016/S0167-7012(00)00218-9
|
| [30] |
Freese HM, Karsten U, Schumann R (2006) Bacterial abundance, activity, and viability in the eutrophic river Warnow, Northeast Germany. Microb Ecol 51: 117–127. doi: 10.1007/s00248-005-0091-5
|
| [31] | Sieracki ME, Cucci TL, Nicinski J (1999) Flow cytometric analysis of 5-cyano-2,3-ditolyl tetrazolium chloride activity of marine bacterioplankton in dilution cultures. Appl Environ Microbiol 65: 2409–2417. |
| [32] |
Mishra A, Taneja N, Sharma M (2012) Viability kinetics, induction, resuscitation and quantitative real-time polymerase chain reaction analyses of viable but nonculturable Vibrio cholerae O1 in freshwater microcosm. J Appl Microbiol 112: 945–953. doi: 10.1111/j.1365-2672.2012.05255.x
|
| [33] | Créach V, Baudoux AC, Bertru G, et al. (2003) Direct estimate of active bacteria: CTC use and limitations. J Microbiol Meth 52: 19–28. |
| [34] |
Purkamo L, Bomberg M, Kietäväinen R, et al. (2016) Microbial co-occurrence patterns in deep Precambrian bedrock fracture fluids. Biogeosciences 13: 3091–3108. doi: 10.5194/bg-13-3091-2016
|
| [35] | Sohlberg E, Bomberg M, Miettinen H, et al., (2015) Revealing the unexplored fungal communities in deep groundwater of crystalline bedrock fracture zones in Olkiluoto, Finland. Front Microbiol 6: 573. |
| [36] | Miettinen H, Kietäväinen R, Sohlberg E, et al. (2015) Microbiome composition and geochemical characteristics of deep subsurface high-pressure environment, Pyhäsalmi mine Finland. Front Microbiol 6. |
| [37] |
Dell'Anno A, Cinzia C (2004) Degradation and turnover of extracellular DNA in marine sediments: ecological and methodological considerations. Appl Environ Microbiol 70: 4384–4386. doi: 10.1128/AEM.70.7.4384-4386.2004
|
| [38] |
Bomberg M, Nyyssönen M, Nousiainen A, et al. (2014) Evaluation of molecular techniques in characterization of deep terrestrial biosphere. OJE 4: 468–487. doi: 10.4236/oje.2014.48040
|
| [39] |
Rajala P, Bomberg M, Vepsäläinen M, et al. (2017) Microbial fouling and corrosion of carbon steel in deep anoxic alkaline groundwater. Biofouling 33: 195–209. doi: 10.1080/08927014.2017.1285914
|
| [40] |
Yakimov MM, La CV, Smedile F, et al. (2014) Heterotrophic bicarbonate assimilation is the main process of de novo organic carbon synthesis in hadal zone of the Hellenic Trench, the deepest part of Mediterranean Sea. Environ Microbiol 6: 709–722. doi: 10.1111/1758-2229.12192
|
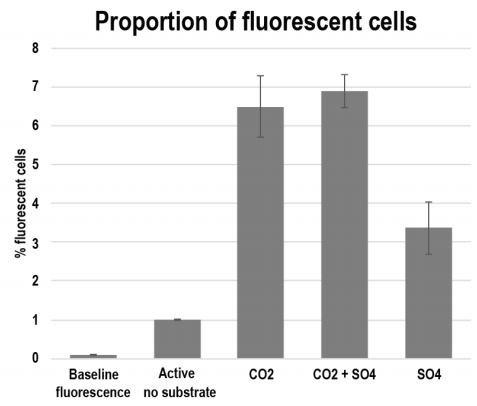
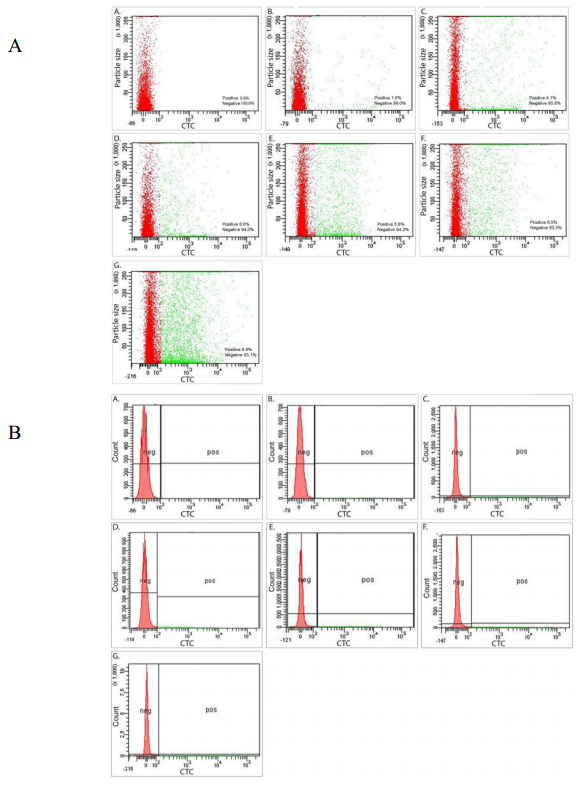
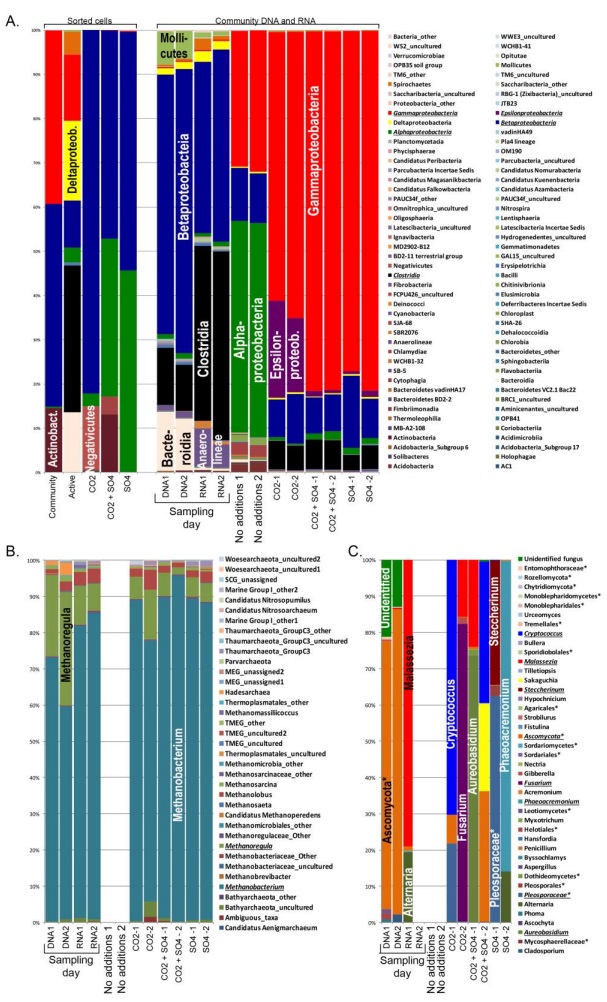
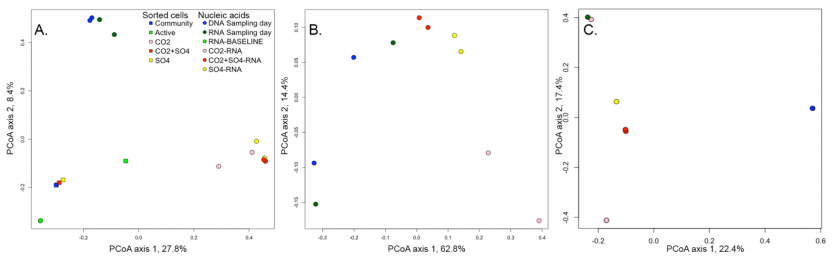
Figures(7) / Tables(2)

Malin Bomberg, Mari Raulio, Sirpa Jylhä, Carsten W. Mueller, Carmen Höschen, Pauliina Rajala, Lotta Purkamo, Riikka Kietäväinen, Lasse Ahonen, Merja Itävaara. CO2 and carbonate as substrate for the activation of the microbial community in 180 m deep bedrock fracture fluid of Outokumpu Deep Drill Hole, Finland[J]. AIMS Microbiology, 2017, 3(4): 846-871. doi: 10.3934/microbiol.2017.4.846







 DownLoad:
DownLoad: