Citation: Sarah Moreland-Russell PhD MPH, Todd Combs PhD, Janice Jones MPH, Amy A. Sorg MPH. State level point-of-sale policy priority as a result of the FSPTCA[J]. AIMS Public Health, 2015, 2(4): 681-690. doi: 10.3934/publichealth.2015.4.681

| [1] | Centers for Disease Control and Prevention (2014) Morbidity and Mortality Weekly Report. 63: 1108-1112. |
| [2] | U.S. Department of Health and Human Services (2014) The Health Consequences of Smoking—50 Years of Progress: A Report of the Surgeon General. Atlanta, GA: Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Office on Smoking and Health. |
| [3] | U.S. Department of Health and Human Services (1994) Preventing Tobacco Use Among Young People: A Report of the Surgeon General. Atlanta, GA: Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Office on Smoking and Health. |
| [4] | U.S. Department of Health and Human Services (2012) Preventing Tobacco Use Among Youth and Young Adults: A Report of the Surgeon General. Atlanta, GA: Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Office on Smoking and Health. |
| [5] | Centers for Disease Control and Prevention (2014) Best Practices for Comprehensive Tobacco Control Programs-2014. Atlanta: U.S. Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Chronic Disease Prevention and Health Promotion, Office on Smoking and Health. |
| [6] | Federal Trade Commission (2015) Cigarette Report for 2012. Federal Trade Commission. |
| [7] | National Cancer Institute (2008) The role of the media in promoting and reducing tobacco use. Bethesda, MD: National Institutes of Health. |
| [8] | Paynter J, Edwards R (2009) The impact of tobacco promotion at the point of sale: A systematic review. Nicotine & Tobacco Res 11: 25-35. |
| [9] |
Slater SJ, Chaloupka FJ, Wakefield M, et al. (2007) The impact of retail cigarette marketing practices on youth smoking uptake. Arch Pediatr Adolesc Med 161: 440-445. doi: 10.1001/archpedi.161.5.440

|
| [10] | San Francisco Board of Supervisors (2014) Ordinance No. 259-14. |
| [11] | Center for Public Health Systems Science (2014) Regulating Pharmacy Tobacco Sales: Massachusetts. St. Louis: Center for Public Health Systems Science, Washington University in St. Louis. |
| [12] |
Luke DA, Ribisl KM, Smith C, et al. (2011) Family Smoking Prevention And Tobacco Control Act: banning outdoor tobacco advertising near schools and playgrounds. American Journal of Preve Med 40: 295-302. doi: 10.1016/j.amepre.2010.11.018
|
| [13] | Ribisl K.M. Luke, D.A., Sorg, A.A., (2011) Reducing tobacco related disparities through point-of-sale regulation: differential impact of regulating tobacco advertising and sales near schools. Annual Meeting and Convention of the American Public Health Association. Washington, D.C. |
| [14] |
Wakefield M, Germain D, Henriksen L (2008) The effect of retail cigarette pack displays on impulse purchase. Addiction 103: 322-328. doi: 10.1111/j.1360-0443.2007.02062.x
|
| [15] | World Health Organization (2008) MPOWER: A policy package to reverse the tobacco epidemic. Geneva: World Health Organization. |
| [16] | Center for Public Health Systems Science (2014) Point of Sale Strategies: A Tobacco Control Guide. St. Louis, MO: Center for Public Health Systems Science, George Warren Brown School of Social Work at Washington University in St. Louis and the Tobacco Control Legal Consortium. |
| [17] | The Center for Public Health and Tobacco Policy (2010) Tobacco Product Display Restrictions. Boston, MA: The Center for Public Health and Tobacco Policy, New England Law. |
| [18] | Center for Public Health & Tobacco Policy (n.d.) Point of sale display regulation. |
| [19] | Institute for Global Tobacco Control (2013) State of evidence review. |
| [20] | World Health Organization (2013) Parties to the WHO Framework Convention on Tobacco Control. |
| [21] |
Li L, Borland R, Fong G (2013) Impact of point-of-sale tobacco display bans: findings from the International Tobacco Control four Country Survey. Health Edu Res 28: 898-910. doi: 10.1093/her/cyt058
|
| [22] | State Tobacco Control Program Staff (2012 & 2014) Point-of-sale policy activity interviews. In: Center for Public Health Systems Science, editor. |
| [23] | QSR International (2012) NVivo qualitative data analysis software, Version 10. |
| [24] | R Core Team (2013) R: A language and environment for statistical computing.In: Computing RFfS, editor. Vienna, Austria. |
| [25] | American Lung Association (2012) State of tobacco control report 2012. Washington D.C & New York: American Lung Association. |
| [26] | American Lung Association (2015) State of tobacco control report 2014. Washington D.C & New York: American Lung Association. |
| [27] | United States Census Bureau (2015) State Government Tax Collections. |
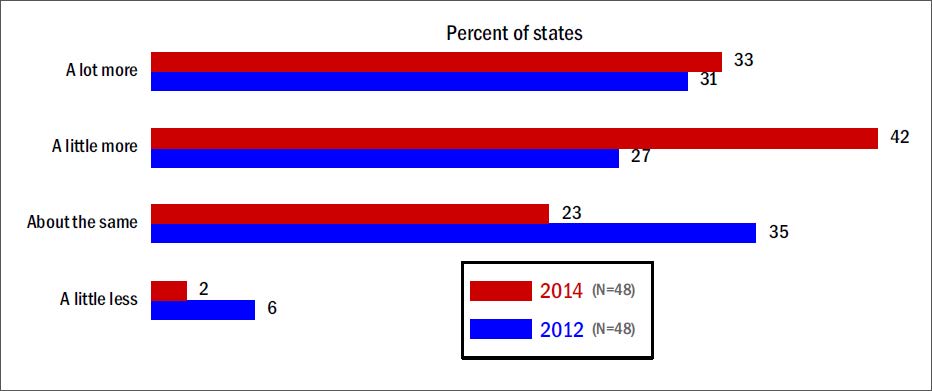
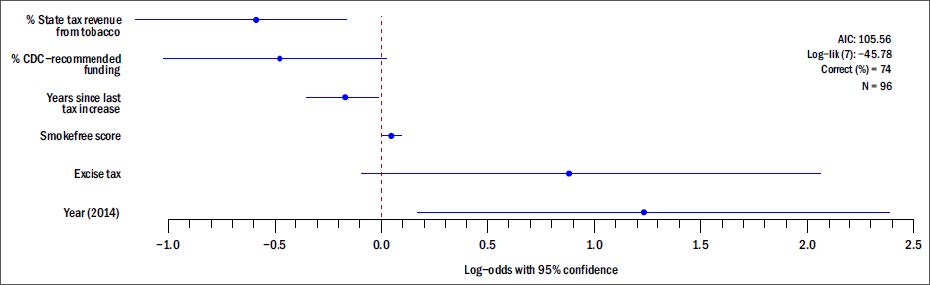
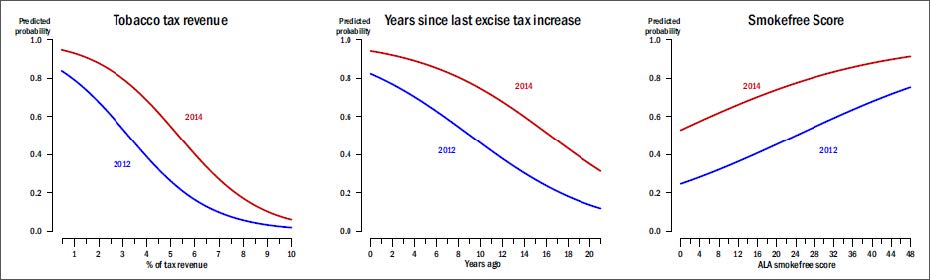
Figures(3) / Tables(2)

Sarah Moreland-Russell PhD MPH, Todd Combs PhD, Janice Jones MPH, Amy A. Sorg MPH. State level point-of-sale policy priority as a result of the FSPTCA[J]. AIMS Public Health, 2015, 2(4): 681-690. doi: 10.3934/publichealth.2015.4.681







 DownLoad:
DownLoad: