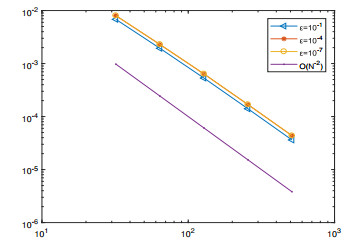
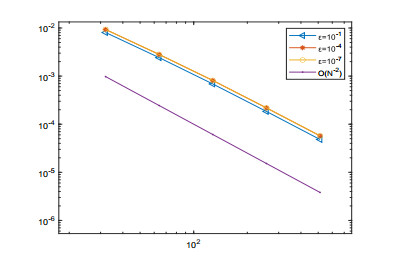
A singularly perturbed Volterra integro-differential problem is considered. The variable two-step backward differentiation formula is used to approximate the first-order derivative term and the trapezoidal formula is used to discretize the integral term. Then, the stability and convergence analysis of the proposed numerical method are proved. It is shown that the proposed scheme is second-order uniformly convergent with respect to perturbation parameter $ \varepsilon $ in the discrete maximum norm. Finally, a numerical experiment verifies the theoretical results.
Citation: Li-Bin Liu, Yige Liao, Guangqing Long. Error estimate of BDF2 scheme on a Bakhvalov-type mesh for a singularly perturbed Volterra integro-differential equation[J]. Networks and Heterogeneous Media, 2023, 18(2): 547-561. doi: 10.3934/nhm.2023023
A singularly perturbed Volterra integro-differential problem is considered. The variable two-step backward differentiation formula is used to approximate the first-order derivative term and the trapezoidal formula is used to discretize the integral term. Then, the stability and convergence analysis of the proposed numerical method are proved. It is shown that the proposed scheme is second-order uniformly convergent with respect to perturbation parameter $ \varepsilon $ in the discrete maximum norm. Finally, a numerical experiment verifies the theoretical results.

| [1] |
G. A. Bocharov, F. A. Rihan, Numerical modelling in biosciences using delay differential equations, J. Comput. Appl. Math., 125 (2000), 183–199. https://doi.org/10.1016/s0377-0427(00)00468-4 doi: 10.1016/s0377-0427(00)00468-4

|
| [2] |
A. De Gaetano, O. Arino, Mathematical modelling of the intravenous glucose tolerance test, J. Math. Biol., 40 (2000), 136–168. https://doi.org/10.1007/s002850050007 doi: 10.1007/s002850050007
|
| [3] |
S. Marino, E. Beretta, D. E. Kirschner, The role of delays in innate and adaptive immunity to intracellular bacterial infection, Math. Biosci. Eng., 4 (2007), 261–286. https://doi.org/10.3934/mbe.2007.4.261 doi: 10.3934/mbe.2007.4.261
|
| [4] |
B. C. Iragi, J. B. Munyakazi, A uniformly convergent numerical method for a singularly perturbed Volterra integro-differential equation, Int. J. Comput. Math., 97 (2020), 759–771. https://doi.org/10.1080/00207160.2019.1585828 doi: 10.1080/00207160.2019.1585828
|
| [5] |
N. N. Nefedov, A. G. Nikitin, T. A. Urazgil'dina, The Cauchy problem for a singularly perturbed Volterra integro-differential equation, Comput. Math. and Math. Phys., 46 (2006), 768–775. https://doi.org/10.1134/s0965542506050046 doi: 10.1134/s0965542506050046
|
| [6] |
A. A. Salama, S. A. Bakr, Difference schemes of exponential type for singularly perturbed Volterra integro-differential problems, Appl. Math. Model., 31 (2007), 866–879. https://doi.org/10.1016/j.apm.2006.02.007 doi: 10.1016/j.apm.2006.02.007
|
| [7] | J. I. Ramos, Exponential techniques and implicit Runge-Kutta method for singularly-perturbed Volterra integro-differential equations, Neural Parallel Sci. Comput., 16 (2008), 387–404. |
| [8] |
S. Şevgin, Numerical solution of a singularly perturbed Volterra integro-differential equation, Adv. Differ. Equ., 2014 (2014), 171. https://doi.org/10.1186/1687-1847-2014-171 doi: 10.1186/1687-1847-2014-171
|
| [9] |
J. Huang, Z. Cen, A. Xu, L. B. Liu, A posteriori error estimation for a singularly perturbed Volterra integro-differential equation, Numer. Algor., 83 (2020), 549–563. https://doi.org/10.1007/s11075-019-00693-y doi: 10.1007/s11075-019-00693-y
|
| [10] |
Sumit, S. Kumar, J. Vigo-Aguiar, Analysis of a nonlinear singularly perturbed Volterra integro-differential equation, J. Comput. Appl. Math., 404 (2021), 113410. https://doi.org/10.1016/j.cam.2021.113410 doi: 10.1016/j.cam.2021.113410
|
| [11] |
X. M. Gu, S. L. Wu, A parallel-in-time iterative algorithm for Volterra partial integro-differential problems with weakly singular kernel, J. Comput. Phys., 417 (2020), 109576. https://doi.org/10.1016/j.jcp.2020.109576 doi: 10.1016/j.jcp.2020.109576
|
| [12] |
Y. L. Zhao, X. M. Gu, A. Ostermann, A preconditioning technique for an all-at-once system from Volterra subdiffusion equations with graded time steps, J. Sci. Comput., 88 (2021), 11. https://doi.org/10.1007/s10915-021-01527-7 doi: 10.1007/s10915-021-01527-7
|
| [13] |
G. Long, L. B. Liu, Z. Huang, Richardson extrapolation method on an adaptive grid for singularly perturbed Volterra integro-differential equations, Numer. Funct. Anal. Optim., 42 (2021), 739–757. https://doi.org/10.1080/01630563.2021.1928698 doi: 10.1080/01630563.2021.1928698
|
| [14] |
O. Yapman, G.M. Amiraliyev, A novel second-order fitted computational method for a singularly perturbed Volterra integro-differential equation, Int. J. Comput. Math., 97 (2020), 1293–1302. https://doi.org/10.1080/00207160.2019.1614565 doi: 10.1080/00207160.2019.1614565
|
| [15] |
W. Chen, X. Wang, Y. Yan, Z. Zhang, A second order BDF numerical scheme with variable steps for the Cahn-Hilliard equation, SIAM J. Numer. Anal., 57 (2019), 495–525. https://doi.org/10.1137/18m1206084 doi: 10.1137/18m1206084
|
| [16] |
E. Emmrich, Stability and error of the variable two-step BDF for semilinear parabolic problems, J. Appl. Math. Computing., 19 (2005), 33–55. https://doi.org/10.1007/bf02935787 doi: 10.1007/bf02935787
|
| [17] |
H. L. Liao, Z. Zhang, Analysis of adaptive BDF2 scheme for diffusion equations, Math. Comp., 90 (2020), 1207–1226. https://doi.org/10.1090/mcom/3585 doi: 10.1090/mcom/3585
|
| [18] |
L. B. Liu, Y. Liao, G. Long, A novel parameter-uniform numerical method for a singularly perturbed Volterra integro-differential equation, Comp. Appl. Math., 42 (2023), 12. https://doi.org/10.1007/s40314-022-02142-4 doi: 10.1007/s40314-022-02142-4
|
| [19] |
X. M. Gu, Y. L. Zhao, X. L. Zhao, C. Bruno; Y. Y. Huang, A note on parallel preconditioning for the all-at-once solution of Riesz fractional diffusion equations, Numer. Math. Theor. Meth. Appl., 14 (2021), 893–919. https://doi.org/10.4208/nmtma.OA-2020-0020 doi: 10.4208/nmtma.OA-2020-0020
|
| [20] |
H. G. Roos, Error estimates for linear finite elements on Bakhvalov-type meshes, Appl Math, 51 (2006), 63–72. https://doi.org/10.1007/s10492-006-0005-y doi: 10.1007/s10492-006-0005-y
|
| [21] |
H. G. Roos, M. Schopf, Analysis of finite element methods on Bakhvalov-type meshes for linear convection-diffusion problems in 2D, Appl. Math., 57 (2012), 97–108. https://doi.org/10.1007/s10492-012-0007-x doi: 10.1007/s10492-012-0007-x
|
| [22] | T. A. Nhan, A uniform convergence analysis for a Bakhvalov-type mesh with an explicitly defined transition point, Numerical Geometry, Grid Generation and Scientific Computing, New York City: Springer, 2021,136–168. |
| [23] |
H. G. Roos, T. Linß, Sufficient conditions for uniform convergence on layer-adapted grids, Computing, 63 (1999), 27–45. https://doi.org/10.1007/s006070050049 doi: 10.1007/s006070050049
|
| [24] | T. Linß, Sufficient conditions for uniform convergence on layer-adapted grids, Appl. Numer. Math., 37 (2001), 241–255. |
| [25] |
C. Li, Q. Yi, A. Chen, Finite difference methods with non-uniform meshes for nonlinear fractional differential equations, J. Comput. Phys., 316 (2016), 614–631. https://doi.org/10.1016/j.jcp.2016.04.039 doi: 10.1016/j.jcp.2016.04.039
|
| [26] |
T. Linß, Error expansion for a first-order upwind difference scheme applied to a model convection-diffusion problem, IMA J. Numer. Anal., 24 (2004), 239–253. https://doi.org/10.1093/imanum/24.2.239 doi: 10.1093/imanum/24.2.239
|
Figures(2) / Tables(2)
Li-Bin Liu, Yige Liao, Guangqing Long. Error estimate of BDF2 scheme on a Bakhvalov-type mesh for a singularly perturbed Volterra integro-differential equation[J]. Networks and Heterogeneous Media, 2023, 18(2): 547-561. doi: 10.3934/nhm.2023023







 DownLoad:
DownLoad: