Cellular Automata have been successfully used to model evolution of complex systems based on simples rules. In this paper we introduce controlled cellular automata to depict the dynamics of systems with controls that can affect their evolution. Using theory from discrete control systems, we derive results for the control of cellular automata in specific cases. The paper is mostly oriented toward two applications: fire spreading; morphogenesis and tumor growth. In both cases, we illustrate the impact of a control on the evolution of the system. For the fire, the control is assumed to be either firelines or firebreaks to prevent spreading or dumping of water, fire retardant and chemicals (foam) on the fire to neutralize it. In the case of cellular growth, the control describes mechanisms used to regulate growth factors and morphogenic events based on the existence of extracellular matrix structures called fractones. The hypothesis is that fractone distribution may coordinate the timing and location of neural cell proliferation, thereby guiding morphogenesis, at several stages of early brain development.
Citation: Achilles Beros, Monique Chyba, Oleksandr Markovichenko. Controlled cellular automata[J]. Networks and Heterogeneous Media, 2019, 14(1): 1-22. doi: 10.3934/nhm.2019001
Cellular Automata have been successfully used to model evolution of complex systems based on simples rules. In this paper we introduce controlled cellular automata to depict the dynamics of systems with controls that can affect their evolution. Using theory from discrete control systems, we derive results for the control of cellular automata in specific cases. The paper is mostly oriented toward two applications: fire spreading; morphogenesis and tumor growth. In both cases, we illustrate the impact of a control on the evolution of the system. For the fire, the control is assumed to be either firelines or firebreaks to prevent spreading or dumping of water, fire retardant and chemicals (foam) on the fire to neutralize it. In the case of cellular growth, the control describes mechanisms used to regulate growth factors and morphogenic events based on the existence of extracellular matrix structures called fractones. The hypothesis is that fractone distribution may coordinate the timing and location of neural cell proliferation, thereby guiding morphogenesis, at several stages of early brain development.

| [1] |
A cellular automaton model for tumour growth in inhomogeneous environment. Journal of Theoretical Biology (2003) 225: 257-274. 
|
| [2] |
A. Beros, M. Chyba, A. Fronville and F. Mercier, A morphogenetic cellular automaton, in 2018 Annual American Control Conference (ACC), IEEE, 2018, 1987–1992. |
| [3] |
A. B. Bishop, Introduction to Discrete Linear Controls: Theory and Application, Elsevier, 2014. |
| [4] |
Cell-level finite element studies of viscous cells in planar aggregates. Journal of Biomechanical Engineering (2000) 122: 394-401.
|
| [5] |
Fractone-heparan sulphates mediate fgf-2 stimulation of cell proliferation in the adult subventricular zone. Cell Proliferation (2013) 46: 137-145.
|
| [6] |
S. El Yacoubi and P. Jacewicz, Cellular automata and controllability problem, in CD-Rom Proceeding of the 14th Int. Symp. on Mathematical Theory of Networks and Systems, june, 2000, 19–23. |
| [7] | Analyse et contrôle par automates cellulaires. Annals of the University of Craiova-Mathematics and Computer Science Series (2003) 30: 210-221. |
| [8] |
Novel extracellular matrix structures in the neural stem cell niche capture the neurogenic factor fibroblast growth factor 2 from the extracellular milieu. Stem Cells (2007) 25: 2146-2157.
|
| [9] |
Emergence and control of macro-spatial structures in perturbed cellular automata, and implications for pervasive computing systems. IEEE Transactions on Systems, Man, and Cybernetics-Part A: Systems and Humans (2005) 35: 337-348.
|
| [10] |
Fractones: Extracellular matrix niche controlling stem cell fate and growth factor activity in the brain in health and disease. Cellular and Molecular Life Sciences (2016) 73: 4661-4674.
|
| [11] |
Bone morphogenetic protein-4 inhibits adult neurogenesis and is regulated by fractone-associated heparan sulfates in the subventricular zone. Journal of Chemical Neuroanatomy (2014) 57: 54-61.
|
| [12] |
Anatomy of the brain neurogenic zones revisited: Fractones and the fibroblast/macrophage network. Journal of Comparative Neurology (2002) 451: 170-188.
|
| [13] | Adhesion between cells, diffusion of growth factors, and elasticity of the aer produce the paddle shape of the chick limb. Physica A: Statistical Mechanics and its Applications (2007) 373: 521-532. |
| [14] |
An integrated agent-mathematical model of the effect of intercellular signalling via the epidermal growth factor receptor on cell proliferation. Journal of Theoretical Biology (2006) 242: 774-789.
|





Figures(14) / Tables(1)
Achilles Beros, Monique Chyba, Oleksandr Markovichenko. Controlled cellular automata[J]. Networks and Heterogeneous Media, 2019, 14(1): 1-22. doi: 10.3934/nhm.2019001

Left: Configuration
Let
In all the figures, red represents positive values and blue represents negative values. The uncontrolled simulation at 4 different timesteps: (a) 0, (b) 5, (c) 20 and (d) 100. The average value at the four timesteps: (a) 0.010, (b) 0.0079, (c) 0.0041 and (d) 0.00014
The pictures show a controlled simulation at the following timesteps: (a) 0, (b) 5, (c) 20, (d) 39, (e) 40, (f) 50, (g) 75, (h) 100 and (i) 150. The control switches on at 20, off at 40, back on at 50 and finally off again at 100. Notice that while the control is off, the sign is constant and the magnitude diminishes at a slow exponential rate. When the control is on, the sign alternates with each timestep and the magnitude increases at a slow exponential rate
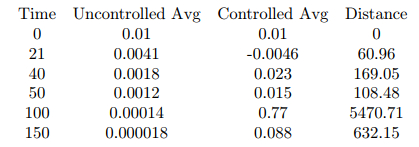
The average is negative for the odd numbered timesteps for which the control is active. For reference, the distance from the controlled simulation at timestep 150 to a grid of zeros is 632.06
Top row using the Von Neumann neighborhood. Timesteps: (a) 1, (b) 50 and (c) 100. Bottom row using the Moore neighborhood. Timesteps: (a) 1, (b) 25 and (c) 50. Note that the Moore neighborhood promotes much faster evolution. For both
Top row:
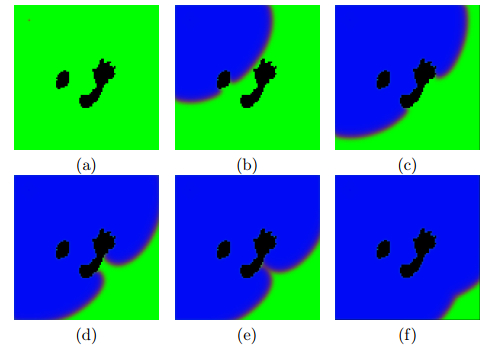
Timesteps: (a) 1, (b) 125, (c) 175, (d) 200, (e) 210 and (f) 250. The fire is diverted by the obstacles
Timesteps: (a) 1, (b) 20, (c) 30, (d) 38, (e) 45 and (f) 100. The fire is diverted by the obstacles
Timesteps: (a) 1, (b) 20, (c) 25, (d) 35, (e) 50 and (f) 100. The fire is diverted by the obstacles
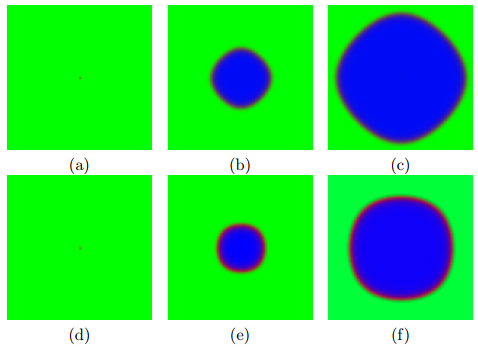
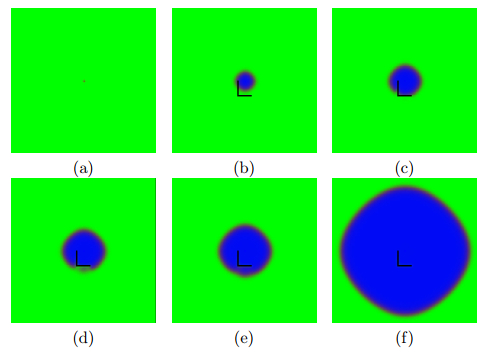
Timesteps: (a) 1, (b) 15, (c) 50 and (d) 100. A simple, uncontrolled cell growth using the Von Neumann neighborhood
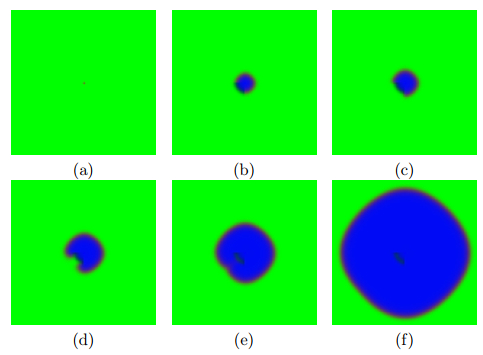
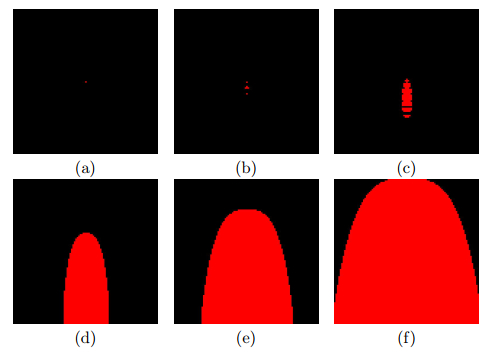
Timesteps: (a) 1, (b) 5, (c) 10, (d) 30, (e) 60 and (f) 100. An uncontrolled cell growth using a neighborhood consisting of the three grid units directly above the central unit and the three side-by-side units in the row three rows below the central unit
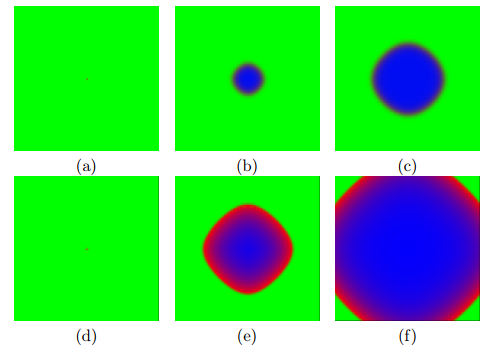
Timesteps: (a) 0, (b) 30, (c) 60 and (d) 100. The interaction between growth of normal cells and tumor cells. The competition between the cell masses plays out at the boundary between the cell masses. The normal cells are in red, the probability of a tumor cell developing is in blue
Timesteps: (a) 1, (b) 5, (c) 15, (d) 30, (e) 60 and (f) 100. Fractones are placed along three horizontal lines. One is above the initial cell and consists of fractones that stop cell growth; they are arranged in blocks. One is in line with the initial cell (across the middle of the simulation) and greatly increases cell growth. The final line is below the intial cell and also stops cell growth. The placement of the fractones is clearly visible in (f)






 DownLoad:
DownLoad: