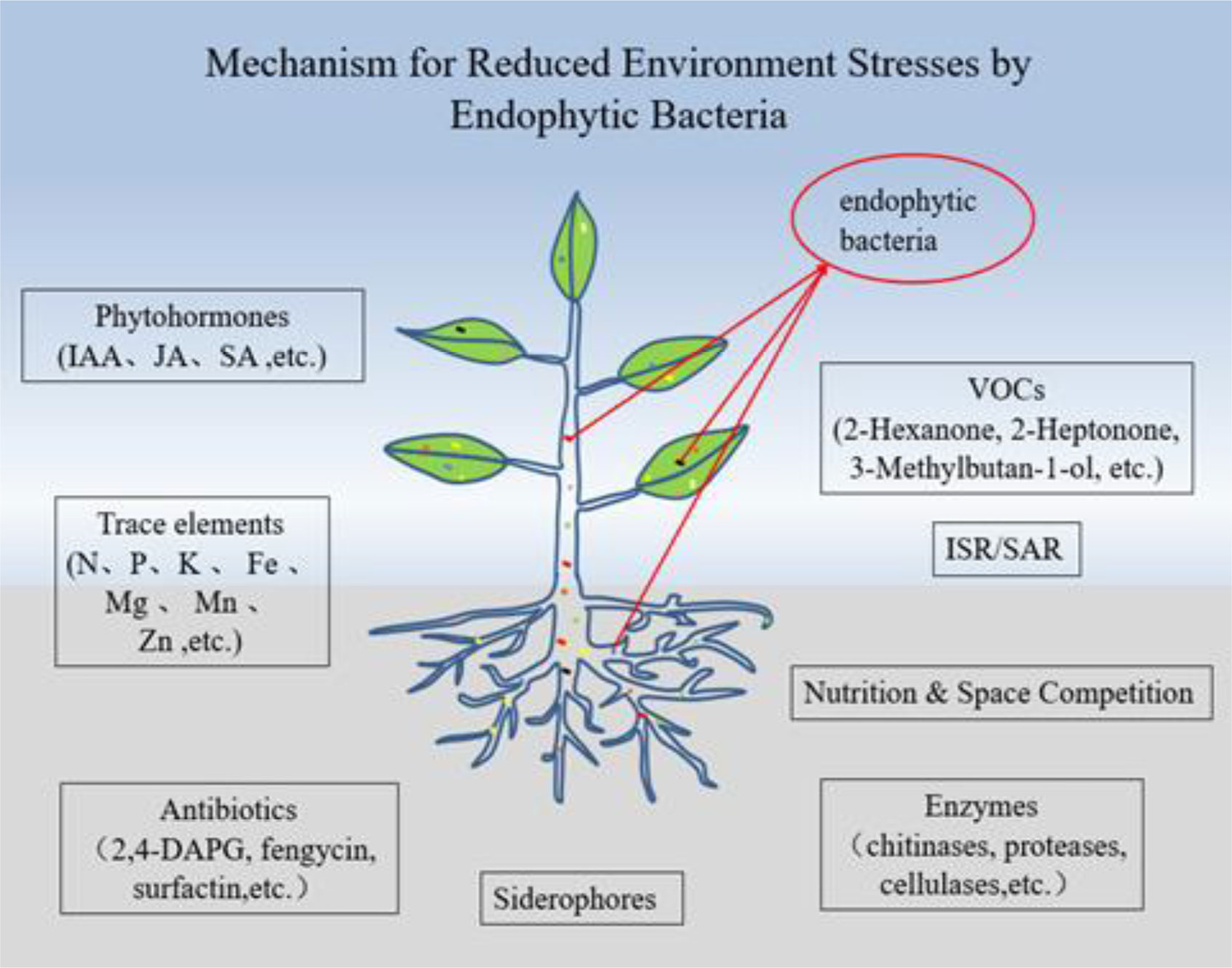
Beneficial endophytic bacteria influence their host plant to grow and resist pathogens. Despite the advantages of endophytic bacteria to their host, their application in agriculture has been low. Furthermore, many plant growers improperly use synthetic chemicals due to having no or little knowledge of the role of endophytic bacteria in plant growth, the prevention and control of pathogens and poor access to endobacterial bioproducts. These synthetic chemicals have caused soil infertility, environmental contamination, disruption to ecological cycles and the emergence of resistant pests and pathogens. There is more that needs to be done to explore alternative ways of achieving sustainable plant production while maintaining environmental health. In recent years, the use of beneficial endophytic bacteria has been noted to be a promising tool in promoting plant growth and the biocontrol of pathogens. Therefore, this review discusses the roles of endophytic bacteria in plant growth and the biocontrol of plant pathogens. Several mechanisms that endophytic bacteria use to alleviate plant biotic and abiotic stresses by helping their host plants acquire nutrients, enhance plant growth and development and suppress pathogens are explained. The review also indicates that there is a gap between research and general field applications of endophytic bacteria and suggests a need for collaborative efforts between growers at all levels. Furthermore, the presence of scientific and regulatory frameworks that promote advanced biotechnological tools and bioinoculants represents major opportunities in the applications of endophytic bacteria. The review provides a basis for future research in areas related to understanding the interactions between plants and beneficial endophytic microorganisms, especially bacteria.
Citation: Rudoviko Galileya Medison, Litao Tan, Milca Banda Medison, Kenani Edward Chiwina. Use of beneficial bacterial endophytes: A practical strategy to achieve sustainable agriculture[J]. AIMS Microbiology, 2022, 8(4): 624-643. doi: 10.3934/microbiol.2022040
Beneficial endophytic bacteria influence their host plant to grow and resist pathogens. Despite the advantages of endophytic bacteria to their host, their application in agriculture has been low. Furthermore, many plant growers improperly use synthetic chemicals due to having no or little knowledge of the role of endophytic bacteria in plant growth, the prevention and control of pathogens and poor access to endobacterial bioproducts. These synthetic chemicals have caused soil infertility, environmental contamination, disruption to ecological cycles and the emergence of resistant pests and pathogens. There is more that needs to be done to explore alternative ways of achieving sustainable plant production while maintaining environmental health. In recent years, the use of beneficial endophytic bacteria has been noted to be a promising tool in promoting plant growth and the biocontrol of pathogens. Therefore, this review discusses the roles of endophytic bacteria in plant growth and the biocontrol of plant pathogens. Several mechanisms that endophytic bacteria use to alleviate plant biotic and abiotic stresses by helping their host plants acquire nutrients, enhance plant growth and development and suppress pathogens are explained. The review also indicates that there is a gap between research and general field applications of endophytic bacteria and suggests a need for collaborative efforts between growers at all levels. Furthermore, the presence of scientific and regulatory frameworks that promote advanced biotechnological tools and bioinoculants represents major opportunities in the applications of endophytic bacteria. The review provides a basis for future research in areas related to understanding the interactions between plants and beneficial endophytic microorganisms, especially bacteria.

| [1] |
Parniske M (2018) Uptake of bacteria into living plant cells, the unifying and distinct feature of the nitrogen-fixing root nodule symbiosis. Curr Opin Plant Biol 44: 164-174. https://doi.org/10.1016/j.pbi.2018.05.016 
|
| [2] | Eid AM, Salim SS, Hassan SED, et al. (2019) Role of endophytes in plant health and abiotic stress management. Microbiome Plant Heal Dis 119–144. https://doi.org/10.1007/978-981-13-8495-0_6 |
| [3] |
Lin H, Liu C, Peng Z, et al. (2022) Distribution pattern of endophytic bacteria and fungi in tea plants. Front Microbiol 13: 872034. https://doi.org/10.3389/fmicb.2022.872034
|
| [4] | Rani L, Pandey RK, Kumar V (2018) Isolation and screening of P-Solubilising endophytic diazotropic bacteria from ethno-medicinal indigenous rice of Jharkhand. Int J Biotechnol Res 5: 1-10. |
| [5] | Tufail MA, Ayyub M, Irfan M, et al. (2022) Endophytic bacteria perform better than endophytic fungi in improving plant growth under drought stress: A meta-comparison spanning 12 years ( 2010 – 2021 ). Physiol Plant 22: e13806. https://doi.org/10.1111/ppl.13806 |
| [6] |
Wang S, Ji B, Su X, et al. (2020) Isolation of endophytic bacteria from Rehmannia glutinosa libosch and their potential to promote plant growth. J Gen Appl Microbiol 66: 279-288. https://doi.org/10.2323/jgam.2019.12.001
|
| [7] | Khan MS, Gao J, Chen X, et al. (2020) Isolation and characterization of plant growth-promoting endophytic bacteria Paenibacillus polymyxa SK1 from Lilium lancifolium. Biomed Res Int 2020: 8650957. https://doi.org/10.1155/2020/8650957 |
| [8] |
Ali S, Isaacson J, Kroner Y, et al. (2018) Corn sap bacterial endophytes and their potential in plant growth-promotion. Environ Sustain 1: 341-355. https://doi.org/10.1007/s42398-018-00030-4
|
| [9] |
Dudeja SS, Suneja-Madan P, Paul M, et al. (2021) Bacterial endophytes: molecular interactions with their hosts. J Basic Microbiol 61: 475-505. https://doi.org/10.1002/jobm.202000657
|
| [10] | Patle P, Phule Krishi Vidyapeeth M, Navnage IN, et al. (2018) Endophytes in plant system: roles in growth promotion, mechanism and their potentiality in achieving agriculture sustainability. Int J Chem Stud 6: 270-274. |
| [11] |
Chen S, Qin R, Yang D, et al. (2022) A comparison of rhizospheric and endophytic bacteria in early and late-maturing pumpkin varieties. Microorganisms 10: 1667. https://doi.org/10.3390/microorganisms10081667
|
| [12] |
White JF, Kingsley KL, Zhang Q, et al. (2019) Review: endophytic microbes and their potential applications in crop management. Pest Manag Sci 75: 2558-2565. https://doi.org/10.1002/ps.5527
|
| [13] |
Hong CE, Park JM (2016) Endophytic bacteria as biocontrol agents against plant pathogens: current state-of-the-art. Plant Biotechnol Rep 10: 353-357. https://doi.org/10.1007/s11816-016-0423-6
|
| [14] | Tavarideh F, Pourahmad F, Nemati M (2022) Diversity and antibacterial activity of endophytic bacteria associated with medicinal plant, Scrophularia striata. Vet Res Forum 13: 409-415. https://doi.org/10.30466/vrf.2021.529714.3174 |
| [15] |
de Carvalho EX, Menezes RSC, de Freitas ADS, et al. (2017) The 15N natural abundance technique to assess the potential of biological nitrogen fixation (BNF) in some important C4 grasses. Aust J Crop Sci 11: 1559-1564. https://doi.org/10.21475/ajcs.17.11.12.pne729
|
| [16] |
Afzal I, Shinwari ZK, Sikandar S, et al. (2019) Plant beneficial endophytic bacteria: mechanisms, diversity, host range and genetic determinants. Microbiol Res 221: 36-49. https://doi.org/10.1016/j.micres.2019.02.001
|
| [17] |
Baldani JI, Reis VM, Videira SS, et al. (2014) The art of isolating nitrogen-fixing bacteria from non-leguminous plants using N-free semi-solid media: a practical guide for microbiologists. Plant Soil 384: 413-431. https://doi.org/10.1007/s11104-014-2186-6
|
| [18] | Puri A, Padda KP, Chanway CP (2018) Nitrogen-fixation by endophytic bacteria in agricultural crops: recent advances. Nitrogen in Agriculture - Updates : 73-94. |
| [19] |
Thakur D, Kaushal R, Shyam V (2014) Phosphate solubilising microorganisms: role in phosphorus nutrition of crop plants-a review. Agric Rev 35: 159. https://doi.org/10.5958/0976-0741.2014.00903.9
|
| [20] |
Bashan Y, Kamnev AA, de-Bashan LE (2013) A proposal for isolating and testing phosphate-solubilizing bacteria that enhance plant growth. Biol Fertil Soils 49: 1-2. https://doi.org/10.1007/s00374-012-0756-4
|
| [21] |
Singh YD, Singh MC (2020) Biotechnological aspects of mangrove microorganisms. Biotechnological Utilization of Mangrove Resources 381-398.
|
| [22] | Sigurbjörnsdóttir MA, Andrésson ÓS, Vilhelmsson O (2015) Analysis of the peltigera membranacea metagenome indicates that lichen-associated bacteria are involved in phosphate solubilization. Microbiol (United Kingdom) 161: 989-996. https://doi.org/10.1099/mic.0.000069 |
| [23] |
Etesami H, Emami S, Alikhani HA (2017) Potassium solubilizing bacteria (KSB): mechanisms, promotion of plant growth, and future prospects - a review. J Soil Sci Plant Nutr 17: 897-911. https://doi.org/10.4067/S0718-95162017000400005
|
| [24] |
Yuan ZS, Liu F, Zhang GF (2015) Characteristics and biodiversity of endophytic phosphorus- and potassiumsolubilizing bacteria in moso bamboo (Phyllostachys edulis). Acta Biol Hung 66: 449-459. https://doi.org/10.1556/018.66.2015.4.9
|
| [25] | Kour D, Rana KL, Kaur T, et al. (2020) Potassium solubilizing and mobilizing microbes: biodiversity, mechanisms of solubilization, and biotechnological implication for alleviations of abiotic stress. Trends of Microbial Biotechnology for Sustainable Agriculture and Biomedicine Systems: Diversity and Functional Perspectives . Elsevier 177-202. |
| [26] |
Xi J, Qian K, Shan L, et al. (2022) The potential of mineral weathering of halophilic‑endophytic bacteria isolated from Suaeda salsa and Spartina anglica. Arch Microbiol 204: 1-12. https://doi.org/10.1007/s00203-022-03129-9
|
| [27] |
Feng K, Cai Z, Ding T, et al. (2019) Effects of potassium-solubulizing and photosynthetic bacteria on tolerance to salt stress in maize. J Appl Microbiol 126: 1530-1540. https://doi.org/10.1111/jam.14220
|
| [28] |
Pirhadi M, Enayatizamir N, Motamedi H, et al. (2016) Screening of salt tolerant sugarcane endophytic bacteria with potassium and zinc for their solubilizing and antifungal activity. Biosci Biotechnol Res Commun 9: 530-538. https://doi.org/10.21786/bbrc/9.3/28
|
| [29] |
Velivelli SLS, Sessitsch A, Prestwich BD (2014) The role of microbial inoculants in integrated crop management systems. Potato Res 57: 291-309. https://doi.org/10.1007/s11540-014-9278-9
|
| [30] | Goswami D, Thakker JN, Dhandhukia PC (2016) Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food Agric 2: 1-19. https://doi.org/10.1080/23311932.2015.1127500 |
| [31] | Sayyed RZ, Badgujar MD, Sonawane HM, et al. (2005) Production of microbial iron chelators (siderophores) by fluorescent Pseudomonads. Indian J Biotechnol 4: 484-490. |
| [32] |
Rungin S, Indananda C, Suttiviriya P, et al. (2012) Plant growth enhancing effects by a siderophore-producing endophytic streptomycete isolated from a Thai jasmine rice. Antonie Van Leeuwenhoek 102: 463-472. https://doi.org/10.1007/s10482-012-9778-z
|
| [33] |
Saravanan VS, Madhaiyan M, Osborne J, et al. (2008) Ecological occurrence of Gluconacetobacter diazotrophicus and nitrogen-fixing Acetobacteraceae members: their possible role in plant growth promotion. Microb Ecol 55: 130-140. https://doi.org/10.1007/s00248-007-9258-6
|
| [34] |
Kamran S, Shahid I, Baig DN, et al. (2017) Contribution of zinc solubilizing bacteria in growth promotion and zinc content of wheat. Front Microbiol 8: 2593. https://doi.org/10.3389/fmicb.2017.02593
|
| [35] |
Mumtaz MZ, Ahmad M, Jamil M, et al. (2017) Zinc solubilizing Bacillus spp. potential candidates for biofortification in maize. Microbiol Res 202: 51-60. https://doi.org/10.1016/j.micres.2017.06.001
|
| [36] | Vaid SK, Kumar B, Sharma A, et al. (2014) Effect of zinc solubilizing bacteria on growth promotion and zinc nutrition of rice. J Soil Sci Plant Nutr 14: 889-910. https://doi.org/10.4067/s0718-95162014005000071 |
| [37] |
Rehman A, Farooq M, Naveed M, et al. (2018) Seed priming of Zn with endophytic bacteria improves the productivity and grain biofortification of bread wheat. Eur J Agron 94: 98-107. https://doi.org/10.1016/j.eja.2018.01.017
|
| [38] |
Sharma P, Kumawat KCK, Kaur S, et al. (2011) Assessment of zinc solubilization by endophytic bacteria in legume rhizosphere. Indian J Appl Res 4: 439-441. https://doi.org/10.15373/2249555x/june2014/137
|
| [39] |
Ahemad M, Kibret M (2014) Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J King Saud Univ - Sci 26: 1-20. https://doi.org/10.1016/j.jksus.2013.05.001
|
| [40] | Role T, Bacteria PP, Stress AS, et al. (2013) The role of phytohormone producing bacteria in alleviating salt stress in crop plants. Biotechnological Techniques of Stress Tolerance in Plants . Stadium Press LLC 21-39. |
| [41] |
Hoffman MT, Gunatilaka MK, Wijeratne K, et al. (2013) Endohyphal bacterium enhances production of indole-3-acetic acid by a foliar fungal endophyte. PLoS One 8: 31-33. https://doi.org/10.1371/journal.pone.0073132
|
| [42] | Zhou H, Ren ZH, Zu X, et al. (2021) Efficacy of plant growth-promoting bacteria Bacillus cereus YN917 for biocontrol of rice blast. Front Microbiol 12: 1-9. https://doi.org/10.3389/fmicb.2021.684888 |
| [43] |
Cohen AC, Travaglia CN, Bottini R, et al. (2009) Participation of abscisic acid and gibberellins produced by endophytic Azospirillum in the alleviation of drought effects in maize. Botany 87: 455-462. https://doi.org/10.1139/B09-023
|
| [44] | Nelson SK, Steber CM (2016) Gibberellin hormone signal perception: down-regulating DELLA repressors of plant growth and development. Annual Plant Reviews . Wiley. |
| [45] |
Martínez C, Espinosa-Ruiz A, Prat S (2016) Gibberellins and plant vegetative growth. Annual Plant Reviews . Wiley 285-322.
|
| [46] |
Padda KP, Puri A, Chanway CP (2016) Effect of GFP tagging of Paenibacillus polymyxa P2b-2R on its ability to promote growth of canola and tomato seedlings. Biol Fertil Soils 52: 377-387. https://doi.org/10.1007/s00374-015-1083-3
|
| [47] |
Padda KP, Puri A, Chanway C (2019) Endophytic nitrogen fixation - a possible “hidden” source of nitrogen for lodgepole pine trees growing at unreclaimed gravel mining sites. FEMS Microbiol Ecol 95: 1-13. https://doi.org/10.1093/femsec/fiz172
|
| [48] |
Wang M, Bian Z, Shi J, et al. (2020) Effect of the nitrogen-fixing bacterium Pseudomonas protegens CHA0-ΔretS-nif on garlic growth under different field conditions. Ind Crops Prod 145: 111982. https://doi.org/10.1016/j.indcrop.2019.111982
|
| [49] |
Abd-Alla MH, Nafady NA, Bashandy SR, et al. (2019) Mitigation of effect of salt stress on the nodulation, nitrogen fixation and growth of chickpea (Cicer arietinum L.) by triple microbial inoculation. Rhizosphere 10: 100148. https://doi.org/10.1016/j.rhisph.2019.100148
|
| [50] |
Chen J, Zhao G, Wei Y, et al. (2021) Isolation and screening of multifunctional phosphate solubilizing bacteria and its growth-promoting effect on Chinese fir seedlings. Sci Rep 11: 1-13. https://doi.org/10.1038/s41598-021-88635-4
|
| [51] | Abo-koura HA, Bishara MM, Saad MM (2019) Isolation and identification of N2 - fixing , phosphate and potassium solubilizing rhizobacteria and their effect on root colonization of wheat plant. Int J Microbiol Res 10: 62-76. https://doi.org/10.5829/idosi.ijmr.2019.62.76 |
| [52] |
Varga T, Hixson KK, Ahkami AH, et al. (2020) Endophyte-promoted phosphorus solubilization in Populus. Front Plant Sci 11: 567918. https://doi.org/10.3389/fpls.2020.567918
|
| [53] |
Mei C, Chretien RL, Amaradasa BS, et al. (2021) Characterization of phosphate solubilizing bacterial endophytes and plant growth promotion in vitro and in greenhouse. Microorganisms 9: 1935. https://doi.org/10.3390/microorganisms9091935
|
| [54] |
Paul D, Sinha SN (2017) Isolation and characterization of phosphate solubilizing bacterium Pseudomonas aeruginosa KUPSB12 with antibacterial potential from river Ganga, India. Ann Agrar Sci 15: 130-136. https://doi.org/10.1016/j.aasci.2016.10.001
|
| [55] |
Baghel V, Thakur JK, Yadav SS, et al. (2020) Phosphorus and potassium solubilization from rock minerals by endophytic Burkholderia sp. strain fdn2-1 in soil and shift in diversity of bacterial endophytes of corn root tissue with crop growth stage. Geomicrobiol J 37: 550-563. https://doi.org/10.1080/01490451.2020.1734691
|
| [56] |
Kushwaha P, Kashyap PL, Srivastava AK, et al. (2020) Plant growth promoting and antifungal activity in endophytic Bacillus strains from pearl millet (Pennisetum glaucum). Brazilian J Microbiol 51: 229-241. https://doi.org/10.1007/s42770-019-00172-5
|
| [57] |
Singh D, Geat N, Rajawat MVS, et al. (2018) Deciphering the mechanisms of endophyte-mediated biofortification of Fe and Zn in wheat. J Plant Growth Regul 37: 174-182. https://doi.org/10.1007/s00344-017-9716-4
|
| [58] |
Singh D, Rajawat MVS, Kaushik R, et al. (2017) Beneficial role of endophytes in biofortification of Zn in wheat genotypes varying in nutrient use efficiency grown in soils sufficient and deficient in Zn. Plant Soil 416: 107-116. https://doi.org/10.1007/s11104-017-3189-x
|
| [59] |
Kang S-M, Waqas M, Hamayun M, et al. (2017) Gibberellins and indole-3-acetic acid producing rhizospheric bacterium Leifsonia xyli SE134 mitigates the adverse effects of copper-mediated stress on tomato. J Plant Interact 12: 373-380. https://doi.org/10.1080/17429145.2017.1370142
|
| [60] |
Dhungana SA, Itoh K (2019) Effects of co-inoculation of indole-3-acetic acid-producing and-degrading bacterial endophytes on plant growth. Horticulturae 5: 17. https://doi.org/10.3390/horticulturae5010017
|
| [61] | Shahzad R, Khan AL, Bilal S, et al. (2017) Plant growth-promoting endophytic bacteria versus pathogenic infections: An example of Bacillus amyloliquefaciens RWL-1 and Fusarium oxysporum f. sp. lycopersici in tomato. PeerJ 2017: 1-21. https://doi.org/10.7717/peerj.3107 |
| [62] |
Kang S, Bilal S, Shahzad R, et al. (2020) Effect of ammonia and indole-3-acetic acid producing endophytic Klebsiella pneumoniae YNA12 as a bio-herbicide for weed inhibition: special reference with evening primroses. Plants (Basel) 9: 761. https://doi.org/10.3390/plants9060761
|
| [63] |
Shah S, Chand K, Rekadwad B, et al. (2021) A prospectus of plant growth promoting endophytic bacterium from orchid (Vanda cristata). BMC Biotechnol 21: 1-9. https://doi.org/10.1186/s12896-021-00676-9
|
| [64] |
Haidar B, Ferdous M, Fatema B, et al. (2018) Population diversity of bacterial endophytes from jute (Corchorus olitorius) and evaluation of their potential role as bioinoculants. Microbiol Res 208: 43-53. https://doi.org/10.1016/j.micres.2018.01.008
|
| [65] |
Khan AL, Halo BA, Elyassi A, et al. (2016) Indole acetic acid and ACC deaminase from endophytic bacteria improves the growth of Solanum lycopersicum. Electron J Biotechnol 21: 58-64. https://doi.org/10.1016/j.ejbt.2016.02.001
|
| [66] |
Hardoim PR, van Overbeek LS, van Elsas JD (2008) Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol 16: 463-471. https://doi.org/10.1016/j.tim.2008.07.008
|
| [67] |
Dell'Amico E, Cavalca L, Andreoni V (2008) Improvement of Brassica napus growth under cadmium stress by cadmium-resistant rhizobacteria. Soil Biol Biochem 40: 74-84. https://doi.org/10.1016/j.soilbio.2007.06.024
|
| [68] |
Sheng XF, Xia JJ, Jiang CY, et al. (2008) Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ Pollut 156: 1164-1170. https://doi.org/10.1016/j.envpol.2008.04.007
|
| [69] |
Wani PA, Khan MS, Zaidi A (2007) Effect of metal tolerant plant growth promoting Bradyrhizobium sp. (vigna) on growth, symbiosis, seed yield and metal uptake by greengram plants. Chemosphere 70: 36-45. https://doi.org/10.1016/j.chemosphere.2007.07.028
|
| [70] |
Mcdonald IR, Murrell JC (1997) The methanol dehydrogenase structural gene mxaf and its use as a functional gene probe for methanotrophs and methylotrophs. Appl Environ Microbiol 63: 3218-3224. https://doi.org/10.1128/aem.63.8.3218-3224.1997
|
| [71] |
Fragner L, Kerou M, Schloter M, et al. (2020) The potato yam phyllosphere ectosymbiont Paraburkholderia sp. Msb3 is a potent growth promotor in tomato. Front Microbio 11: 1-21. https://doi.org/10.3389/fmicb.2020.00581
|
| [72] |
Pieterse CMJ, Zamioudis C, Berendsen RL, et al. (2014) Induced systemic resistance by beneficial microbes. Annu Rev Phytopathol 52: 347-375. https://doi.org/10.1146/annurev-phyto-082712-102340
|
| [73] |
Weller DM, Mavrodi DV, Van Pelt JA, et al. (2012) Induced systemic resistance in Arabidopsis thaliana against Pseudomonas syringae pv. tomato by 2,4-diacetylphloroglucinol-producing Pseudomonas fluorescens. Phytopathology 102: 403-412. https://doi.org/10.1094/PHYTO-08-11-0222
|
| [74] |
Ait Barka E, Gognies S, Nowak J, et al. (2002) Inhibitory effect of endophyte bacteria on Botrytis cinerea and its influence to promote the grapevine growth. Biol Control 24: 135-142. https://doi.org/10.1016/S1049-9644(02)00034-8
|
| [75] |
Wang M, Xing Y, Wang J, et al. (2014) The role of the chi1 gene from the endophytic bacteria Serratia proteamaculans 336x in the biological control of wheat take-all. Can J Microbiol 60: 533-540. https://doi.org/10.1139/cjm-2014-0212
|
| [76] |
Ren J, Li H, Wang Y, et al. (2013) Biocontrol potential of an endophytic Bacillus pumilus JK-SX001 against poplar canker. Biol Control 67: 421-430. https://doi.org/10.1016/j.biocontrol.2013.09.012
|
| [77] |
Cho KM, Hong SY, Lee SM, et al. (2007) Endophytic bacterial communities in ginseng and their antifungal activity against pathogens. Microbial Ecology 54: 341-351. https://doi.org/10.1007/s00248-007-9208-3
|
| [78] |
Gong Q, Zhang C, Lu F, et al. (2014) Identification of bacillomycin D from Bacillus subtilis fmbJ and its inhibition effects against Aspergillus flavus. Food Control 36: 8-14. https://doi.org/10.1016/j.foodcont.2013.07.034
|
| [79] |
Yánez-Mendizábal V, Usall J, Viñas I, et al. (2011) Potential of a new strain of Bacillus subtilis CPA-8 to control the major postharvest diseases of fruit. Biocontrol Sci Technol 21: 409-426. https://doi.org/10.1080/09583157.2010.541554
|
| [80] |
Moyne AL, Shelby R, Cleveland TE, et al. (2001) Bacillomycin D: an iturin with antifungal activity against Aspergillus flavus. J Appl Microbiol 90: 622-629. https://doi.org/10.1046/j.1365-2672.2001.01290.x
|
| [81] |
Martins SJ, Faria AF, Pedroso MP, et al. (2019) Microbial volatiles organic compounds control anthracnose (Colletotrichum lindemuthianum) in common bean (Phaseolus vulgaris L.). Biol Control 131: 36-42. https://doi.org/10.1016/j.biocontrol.2019.01.003
|
| [82] |
Chaouachi M, Marzouk T, Jallouli S, et al. (2021) Activity assessment of tomato endophytic bacteria bioactive compounds for the postharvest biocontrol of Botrytis cinerea. Postharvest Biol Technol 172: 111389. https://doi.org/10.1016/j.postharvbio.2020.111389
|
| [83] |
Jiang CH, Liao MJ, Wang HK, et al. (2018) Bacillus velezensis, a potential and efficient biocontrol agent in control of pepper gray mold caused by Botrytis cinerea. Biol Control 126: 147-157. https://doi.org/10.1016/j.biocontrol.2018.07.017
|
| [84] |
Ryan RP, Germaine K, Franks A, et al. (2008) Bacterial endophytes: recent developments and applications. FEMS Microbiol Lett 278: 1-9. https://doi.org/10.1111/j.1574-6968.2007.00918.x
|
| [85] |
Pathak P, Rai VK, Can H, et al. (2022) Plant-endophyte interaction during biotic stress management. Plants (Basel) 11: 2203. https://doi.org/10.3390/plants1117220
|
| [86] |
Ali M, Ahmad Z, Ashraf MF, et al. (2021) Maize endophytic microbial-communities revealed by removing PCR and 16S rRNA sequencing and their synthetic applications to suppress maize banded leaf and sheath blight. Microbiol Res 242: 126639. https://doi.org/10.1016/j.micres.2020.126639
|
| [87] |
Nawangsih Aa, Damayanti Ika, Wiyono S, et al. (2011) Selection and characterization of endophytic bacteria as biocontrol agents of tomato bacterial wilt disease. HAYATI J Biosci 18: 66-70. https://doi.org/10.4308/hjb.18.2.66
|
| [88] |
Eljounaidi K, Lee SK, Bae H (2016) Bacterial endophytes as potential biocontrol agents of vascular wilt diseases–review and future prospects. Biol Control 103: 62-68. https://doi.org/10.1016/j.biocontrol.2016.07.013
|
| [89] |
Aravind R, Eapen SJ, Kumar A, et al. (2010) Screening of endophytic bacteria and evaluation of selected isolates for suppression of burrowing nematode (Radopholus similis Thorne) using three varieties of black pepper (Piper nigrum L.). Crop Prot 29: 318-324. https://doi.org/10.1016/j.cropro.2009.12.005
|
| [90] |
Aravind R, Kumar A, Eapen SJ, et al. (2009) Endophytic bacterial flora in root and stem tissues of black pepper (Piper nigrum L.) genotype: isolation, identification and evaluation against Phytophthora capsici. Lett Appl Microbiol 48: 58-64. https://doi.org/10.1111/j.1472-765X.2008.02486.x
|
| [91] | Wu Y, Lu C, Qian X, et al. (2009) Diversities within genotypes, bioactivity and biosynthetic genes of endophytic actinomycetes isolated from three pharmaceutical plants. Curr Microbiol 59: 475–482. https://doi.org/10.1007/s00284-009-9463-2 |
| [92] |
Dowarah B, Agarwal H, Krishnatreya DB, et al. (2021) Evaluation of seed associated endophytic bacteria from tolerant chilli cv. Firingi Jolokia for their biocontrol potential against bacterial wilt disease. Microbiol Res 248: 126751. https://doi.org/10.1016/j.micres.2021.126751
|
| [93] |
Semenzato G, Faddetta T, Falsini S, et al. (2022) Endophytic bacteria associated with Origanum heracleoticum L. (Lamiaceae) seeds. Microorganisms 10: 2086. https://doi.org/10.3390/microorganisms10102086
|
| [94] |
Cui L, Yang C, Wei L, et al. (2020) Isolation and identification of an endophytic bacteria Bacillus velezensis 8-4 exhibiting biocontrol activity against potato scab. Biol Control 141: 104156. https://doi.org/10.1016/j.biocontrol.2019.104156
|
| [95] |
Djaya L, Hersanti, Istifadah N, et al. (2019) In vitro study of plant growth promoting rhizobacteria (PGPR)and endophytic bacteria antagonistic to Ralstonia solanacearum formulated with graphite and silica nano particles as a biocontrol delivery system (BDS). Biocatal Agric Biotechnol 19: 101153. https://doi.org/10.1016/j.bcab.2019.101153
|
| [96] |
Marian M, Ohno T, Suzuki H, et al. (2020) A novel strain of endophytic Streptomyces for the biocontrol of strawberry anthracnose caused by Glomerella cingulata. Microbiol Res 234: 126428. https://doi.org/10.1016/j.micres.2020.126428
|
| [97] |
Zheng T, Liu L, Nie Q, et al. (2021) Isolation, identification and biocontrol mechanisms of endophytic bacterium D61-A from Fraxinus hupehensis against Rhizoctonia solani. Biol Control 158: 104621. https://doi.org/10.1016/j.biocontrol.2021.104621
|
| [98] |
Chen L, Shi H, Heng J, et al. (2019) Antimicrobial, plant growth-promoting and genomic properties of the peanut endophyte Bacillus velezensis LDO2. Microbiol Res 218: 41-48. https://doi.org/10.1016/j.micres.2018.10.002
|
| [99] |
Hong CE, Jeong H, Jo SH, et al. (2016) A leaf-inhabiting endophytic bacterium, Rhodococcus sp. KB6, enhances sweet potato resistance to black rot disease caused by Ceratocystis fimbriata. J Microbiol Biotechnol 26: 488-492. https://doi.org/10.4014/jmb.1511.11039
|
| [100] |
Deng Y, Zhu Y, Wang P, et al. (2011) Complete genome sequence of Bacillus subtilis BSn5, an endophytic bacterium of Amorphophallus konjac with antimicrobial activity for the plant pathogen Erwinia carotovora subsp. carotovora. J Bacteriol 193: 2070-2071. https://doi.org/10.1128/JB.00129-11
|
| [101] |
Yu H, Zhang L, Li L, et al. (2010) Recent developments and future prospects of antimicrobial metabolites produced by endophytes. Microbiol Res 165: 437-449. https://doi.org/10.1016/j.micres.2009.11.009
|
| [102] |
Vrublevskaya M, Gharwalova L, Kolouchova I (2019) Biocontrol properties of endophytic bacteria from Vitis vinifera. J Biotechnol 305: S41. https://doi.org/10.1016/j.jbiotec.2019.05.148
|
| [103] |
Stringlis IA, Zamioudis C, Berendsen RL, et al. (2019) Type III secretion system of beneficial rhizobacteria pseudomonas simiae WCS417 and pseudomonas defensor WCS374. Front Microbiol 10: 1-11. https://doi.org/10.3389/fmicb.2019.01631
|
| [104] |
Compant S, Duffy B, Nowak J, et al. (2005) Use of plant growth-promoting bacteria for biocontrol of plant diseases: principles, mechanisms of action, and future prospects. Appl Environ Microbiol 71: 4951-4959. https://doi.org/10.1128/AEM.71.9.4951-4959.2005
|
| [105] |
Lucas JA, García-Cristobal J, Bonilla A, et al. (2014) Beneficial rhizobacteria from rice rhizosphere confers high protection against biotic and abiotic stress inducing systemic resistance in rice seedlings. Plant Physiol Biochem 82: 44-53. https://doi.org/10.1016/j.plaphy.2014.05.007
|
| [106] | Yi HS, Yang JW, Ryu CM (2013) ISR meets SAR outside: additive action of the endophyte Bacillus pumilus INR7 and the chemical inducer, benzothiadiazole, on induced resistance against bacterial spot in field-grown pepper. Front Plant Sci 4: 1-12. https://doi.org/10.3389/fpls.2013.00122 |
| [107] |
Conn VM, Walker AR, Franco CMM (2008) Endophytic actinobacteria induce defense pathways in Arabidopsis thaliana. Mol Plant-Microbe Interac 21: 208-218. https://doi.org/10.1094/MPMI-21-2-0208
|
| [108] |
Pieterse CMJ, Van Der Does D, Zamioudis C, et al. (2012) Hormonal modulation of plant immunity. Annu Rev Cell Dev Biol 28: 489-521. https://doi.org/10.1146/annurev-cellbio-092910-154055
|
| [109] |
Ryu CM, Farag MA, Hu CH, et al. (2004) Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol 134: 1017-1026. https://doi.org/10.1104/pp.103.026583
|
| [110] |
Ryu CM, Murphy JF, Mysore KS, et al. (2004) Plant growth-promoting rhizobacteria systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. Plant J 39: 381-392. https://doi.org/10.1111/j.1365-313X.2004.02142.x
|
| [111] |
Compant S, Reiter B, Sessitsch A, et al. (2005) Endophytic colonization of Vitis vinifera L. by plant growth-promoting bacterium Burkholderia sp. strain PsJN. Appl Environ Microbiol 71: 1685-1693. https://doi.org/10.1128/AEM.71.4.1685-1693.2005
|
| [112] |
Frankowski J, Lorito M, Scala F, et al. (2001) Purification and properties of two chitinolytic enzymes of Serratia plymuthica HRO-C48. Arch Microbiol 176: 421-426. https://doi.org/10.1007/s002030100347
|
| [113] |
Duijff BJ, Gianinazzi-Pearson V, Lemanceau P (1997) Involvement of the outer membrane lipopolysaccharides in the endophytic colonization of tomato roots by biocontrol Pseudomonas fluorescens strain WCS417r. New Phytol 135: 325-334. https://doi.org/10.1046/j.1469-8137.1997.00646.x
|
| [114] | Pliego C, Kamilova F, Lugtenberg B (2011). Bacteria in agrobiology: crop ecosystems, Springer Berlin, Heidelberg |
| [115] |
Ahmad F, Ahmad I, Khan MS (2008) Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol Res 163: 173-181. https://doi.org/10.1016/j.micres.2006.04.001
|
| [116] |
Zhao LF, Xu YJ, Lai XH (2018) Antagonistic endophytic bacteria associated with nodules of soybean (Glycine max L.) and plant growth-promoting properties. Brazilian J Microbiol 49: 269-278. https://doi.org/10.1016/j.bjm.2017.06.007
|
| [117] |
Martinez-Klimova E, Rodríguez-Peña K, Sánchez S (2017) Endophytes as sources of antibiotics. Biochem Pharmacol 134: 1-17. https://doi.org/10.1016/j.bcp.2016.10.010
|
| [118] |
Xu W, Wang F, Zhang M, et al. (2019) Diversity of cultivable endophytic bacteria in mulberry and their potential for antimicrobial and plant growth-promoting activities. Microbiol Res 229: 126328. https://doi.org/10.1016/j.micres.2019.126328
|
| [119] |
Gond SK, Bergen MS, Torres MS, et al. (2015) Endophytic Bacillus spp. produce antifungal lipopeptides and induce host defence gene expression in maize. Microbiol Res 172: 79-87. https://doi.org/10.1016/j.micres.2014.11.004
|
| [120] |
Castillo UF, Strobel GA, Ford EJ, et al. (2002) Munumbicins, wide-spectrum antibiotics produced by Streptomyces NRRL 30562, endophytic on Kennedia nigriscans. Microbiology 148: 2675-2685. https://doi.org/10.1099/00221287-148-9-2675
|
| [121] |
Etminani F, Harighi B, Mozafari AA (2022) Effect of volatile compounds produced by endophytic bacteria on virulence traits of grapevine crown gall pathogen, Agrobacterium tumefaciens. Sci Rep 12: 10510. https://doi.org/10.1038/s41598-022-14864-w
|
| [122] |
Korpi A, Järnberg J, Pasanen AL (2009) Microbial volatile organic compounds. Crit Rev Toxicol 39: 139-193. https://doi.org/10.1080/10408440802291497
|
| [123] |
Cheffi Azabou M, Gharbi Y, Medhioub I, et al. (2020) The endophytic strain Bacillus velezensis OEE1: an efficient biocontrol agent against Verticillium wilt of olive and a potential plant growth promoting bacteria. Biol Control 142: 104168. https://doi.org/10.1016/j.biocontrol.2019.104168
|
| [124] |
Mohamad OAA, Li L, Ma JB, et al. (2018) Evaluation of the antimicrobial activity of endophytic bacterial populations from Chinese traditional medicinal plant licorice and characterization of the bioactive secondary metabolites produced by Bacillus atrophaeus against Verticillium dahliae. Front Microbiol 9: 1-14. https://doi.org/10.3389/fmicb.2018.00924
|
| [125] |
Husson E, Hadad C, Huet G, et al. (2017) The effect of room temperature ionic liquids on the selective biocatalytic hydrolysis of chitin: via sequential or simultaneous strategies. Green Chem 19: 4122-4131. https://doi.org/10.1039/c7gc01471f
|
| [126] | Vaddepalli P, Fulton L, Wieland J, et al. (2017) The cell wall-localized atypical β-1,3 glucanase ZERZAUST controls tissue morphogenesis in Arabidopsis thaliana. Development 144: 2259-2269. https://doi.org/10.1242/dev.152231 |
| [127] |
Friedrich N, Hagedorn M, Soldati-Favre D, et al. (2012) Prison break: pathogens' strategies to egress from host cells. Microbiol Mol Biol Rev 76: 707-720. https://doi.org/10.1128/mmbr.00024-12
|
| [128] |
Mauch F, Mauch-Mani B, Boller T (1988) Antifungal hydrolases in pea tissue. Plant Physiol 88: 936-942. https://doi.org/10.1104/pp.88.3.936
|
| [129] |
Chen C, Cao Z, Li J, et al. (2020) A novel endophytic strain of Lactobacillus plantarum CM-3 with antagonistic activity against Botrytis cinerea on strawberry fruit. Biol Control 148: 104306. https://doi.org/10.1016/j.biocontrol.2020.104306
|
| [130] |
El-Tarabily KA (2006) Rhizosphere-competent isolates of streptomycete and non-streptomycete actinomycetes capable of producing cell-wall-degrading enzymes to control Pythium aphanidermatum damping-off disease of cucumber. Can J Bot 84: 211-222. https://doi.org/10.1139/B05-153
|
| [131] |
Jha Y (2019) Endophytic bacteria mediated anti‑autophagy and induced catalase , β‑1, 3‑glucanases gene in paddy after infection with pathogen Pyricularia grisea. Indian Phytopathol 72: 99-106. https://doi.org/10.1007/s42360-018-00106-5
|
| [132] |
Rojas CM, Senthil-kumar M, Tzin V, et al. (2014) Regulation of primary plant metabolism during plant-pathogen interactions and its contribution to plant defense. Front Plant Sci 5: 1-12. https://doi.org/10.3389/fpls.2014.00017
|
| [133] |
Mysore KS, Ryu C (2004) Nonhost resistance: how much do we know?. Trends Plant Sci 9: 97-104. https://doi.org/10.1016/j.tplants.2003.12.005
|
| [134] | Ngalimat MS, Hata EM, Zulperi D, et al. (2021) Plant growth-promoting bacteria as an emerging tool to manage bacterial rice pathogens. Microorganisms 9: 1-23. https://doi.org/10.3390/microorganisms9040682 |
| [135] |
Tranier MS, Pognant-Gros J, Quiroz R de la C, et al. (2014) Commercial biological control agents targeted against plant-parasitic root-knot nematodes. Brazilian Arch Biol Technol 57: 831-841. https://doi.org/10.1590/S1516-8913201402540
|
| [136] |
Vurukonda SSKP, Giovanardi D, Stefani E (2018) Plant growth promoting and biocontrol activity of Streptomyces spp. As endophytes. Int J Mol Sci 19: 952. https://doi.org/10.3390/ijms19040952
|
Figures(1) / Tables(1)

Rudoviko Galileya Medison, Litao Tan, Milca Banda Medison, Kenani Edward Chiwina. Use of beneficial bacterial endophytes: A practical strategy to achieve sustainable agriculture[J]. AIMS Microbiology, 2022, 8(4): 624-643. doi: 10.3934/microbiol.2022040







 DownLoad:
DownLoad: