Citation: Sugenendran Supramani, Rahayu Ahmad, Zul Ilham, Mohamad Suffian Mohamad Annuar, Anita Klaus, Wan Abd Al Qadr Imad Wan-Mohtar. Optimisation of biomass, exopolysaccharide and intracellular polysaccharide production from the mycelium of an identified Ganoderma lucidum strain QRS 5120 using response surface methodology[J]. AIMS Microbiology, 2019, 5(1): 19-38. doi: 10.3934/microbiol.2019.1.19

| [1] |
Adotey G, Quarcoo A, Holliday JC, et al. (2011) Effect of immunomodulating and antiviral agent of medicinal mushrooms (immune assist 24/7 (TM)) on CD4+T-lymphocyte counts of HIV-infected patients. Int J Med Mushrooms 13: 109–113. doi: 10.1615/IntJMedMushr.v13.i2.20

|
| [2] |
Andoh T, Zhang Q, Yamamoto T, et al. (2010) Inhibitory effects of the methanol extract of Ganoderma lucidum on mosquito allergy-induced itch-associated responses in mice. J Pharmacol Sci 114: 292–297. doi: 10.1254/jphs.10180FP
|
| [3] |
Baig MN, Shahid AA, Ali M (2015) In vitro assessment of extracts of the lingzhi or reishi medicinal mushroom, Ganoderma lucidum (higher basidiomycetes) against different plant pathogenic fungi. Int J Med Mushrooms 17: 407–411. doi: 10.1615/IntJMedMushrooms.v17.i4.90
|
| [4] |
Barbieri A, Quagliariello V, Del Vecchio V, et al. (2017) Anticancer and anti-inflammatory properties of Ganoderma lucidum extract effects on melanoma and triple-negative breast cancer treatment. Nutrients 9: 210. doi: 10.3390/nu9030210
|
| [5] | Batbayar S, Kim MJ, Kim HW (2011) Medicinal mushroom Lingzhi or Reishi, Ganoderma lucidum (W.Curt.:Fr.) P. Karst., beta-glucan induces toll-like receptors and fails to induce inflammatory cytokines in NF-kappa B inhibitor-treated macrophages. Int J Med Mushrooms 13: 213–225. |
| [6] |
Bao XF, Fang JN, Li XY (2001) Structural characterization and immunomodulating activity of a complex glucan from spores of Ganoderma lucidum. Biosci Biotech Bioch 65: 2384–2391. doi: 10.1271/bbb.65.2384
|
| [7] |
Trajkovic LMH, Mijatovic SA, Maksimovic-Ivanic DD, et al. (2009) Anticancer properties of Ganoderma Lucidum methanol extracts in vitro and in vivo. Nutr Cancer 61: 696–707. doi: 10.1080/01635580902898743
|
| [8] |
Wan-Mohtar WAAQ, Young L, Abbott GM, et al. (2016) Antimicrobial properties and cytotoxicity of sulfated (1,3)-beta-D-glucan from the mycelium of the mushroom Ganoderma lucidum. J Microbiol Biotechnol 26: 999–1010. doi: 10.4014/jmb.1510.10018
|
| [9] | Wan-Mohtar WAAQI, Viegelmann C, Klaus A, et al. (2017) Antifungal-demelanizing properties and RAW264.7 macrophages stimulation of glucan sulfate from the mycelium of the mushroom Ganoderma lucidum. Food Sci Biotechnol 26: 159–165. |
| [10] |
Wan-Mohtar WAAQI, Latif NA, Harvey LM, et al. (2016) Production of exopolysaccharide by Ganoderma lucidum in a repeated-batch fermentation. Biocatal Agr Biotechnol 6: 91–101. doi: 10.1016/j.bcab.2016.02.011
|
| [11] |
Viniegra-Gonzalez G, Favela-Torres E, Aguilar CN, et al. (2003) Advantages of fungal enzyme production in solid state over liquid fermentation systems. Biochem Eng J 13: 157–167. doi: 10.1016/S1369-703X(02)00128-6
|
| [12] |
Wan-Mohtar WAAQI, Malek RA, Harvey LM, et al. (2016) Exopolysaccharide production by Ganoderma lucidum immobilised on polyurethane foam in a repeated-batch fermentation. Biocatal Agr Biotechnol 8: 24–31. doi: 10.1016/j.bcab.2016.08.002
|
| [13] |
Espinosa-Ortiz EJ, Rene ER, Pakshirajan K, et al. (2016) Fungal pelleted reactors in wastewater treatment: Applications and perspectives. Chem Eng J 283: 553–571. doi: 10.1016/j.cej.2015.07.068
|
| [14] |
Ubaidillah N, Hafizah N, Abdullah N, et al. (2015) Isolation of the intracellular and extracellular polysaccharides of Ganoderma neojaponicum (Imazeki) and characterization of their immunomodulatory properties. Electron J Biotech 18: 188–195. doi: 10.1016/j.ejbt.2015.03.006
|
| [15] |
Ahmad R, Al-Shorgani NKN, Hamid AA, et al. (2013) Optimization of medium components using response surface methodology (RSM) for mycelium biomass and exopolysaccharide production by Lentinus squarrosulus. Adv Biosci Biotechnol 4: 1079. doi: 10.4236/abb.2013.412144
|
| [16] | Wu FL, Zhang G, Ren A, et al. (2016) The pH-responsive transcription factor PacC regulates mycelial growth, fruiting body development, and ganoderic acid biosynthesis in Ganoderma lucidum. Mycologia 108: 1104–1113. |
| [17] | Stamets P (1983) The mushroom cultivator: a practical guide to growing mushroom at home. Chilton, JS. |
| [18] |
Liao B, Chen X, Han J, et al. (2015) Identification of commercial Ganoderma (Lingzhi) species by ITS2 sequences. Chin Med 10: 22. doi: 10.1186/s13020-015-0056-7
|
| [19] |
Hennicke F, Cheikh-Ali Z, Liebisch T, et al. (2016) Distinguishing commercially grown Ganoderma lucidum from Ganoderma lingzhi from Europe and East Asia on the basis of morphology, molecular phylogeny, and triterpenic acid profiles. Phytochemistry 127: 29–37. doi: 10.1016/j.phytochem.2016.03.012
|
| [20] |
Hou X, Xiao M, Chen SCA, et al. (2016) Sequencer-based capillary gel electrophoresis (SCGE) Targeting the rDNA internal transcribed spacer (ITS) regions for accurate identification of clinically important yeast species. PLoS One 11: e0154385. doi: 10.1371/journal.pone.0154385
|
| [21] | Park HG, Ko HG, Kim SH, et al. (2004) Molecular identification of Asian isolates of medicinal mushroom Hericium erinaceum by phylogenetic analysis of nuclear ITS rDNA. J Microbiol Biotechn 14: 816–821. |
| [22] |
Parnmen S, Sikaphan S, Leudang S, et al. (2016) Molecular identification of poisonous mushrooms using nuclear ITS region and peptide toxins: a retrospective study on fatal cases in Thailand. J Toxicol Sci 41: 65–76. doi: 10.2131/jts.41.65
|
| [23] |
Chang MY, Tsai GJ, Houng JY (2006) Optimization of the medium composition for the submerged culture of Ganoderma lucidum by Taguchi array design and steepest ascent method. Enzyme Microb Tech 38: 407–414. doi: 10.1016/j.enzmictec.2005.06.011
|
| [24] |
Yuan BJ, Chi XY, Zhang RJ (2012) Optimization of exopolysaccharides production from a novel strain of Ganoderma Lucidum Cau5501 in submerged culture. Braz J Microbiol 43: 490–497. doi: 10.1590/S1517-83822012000200009
|

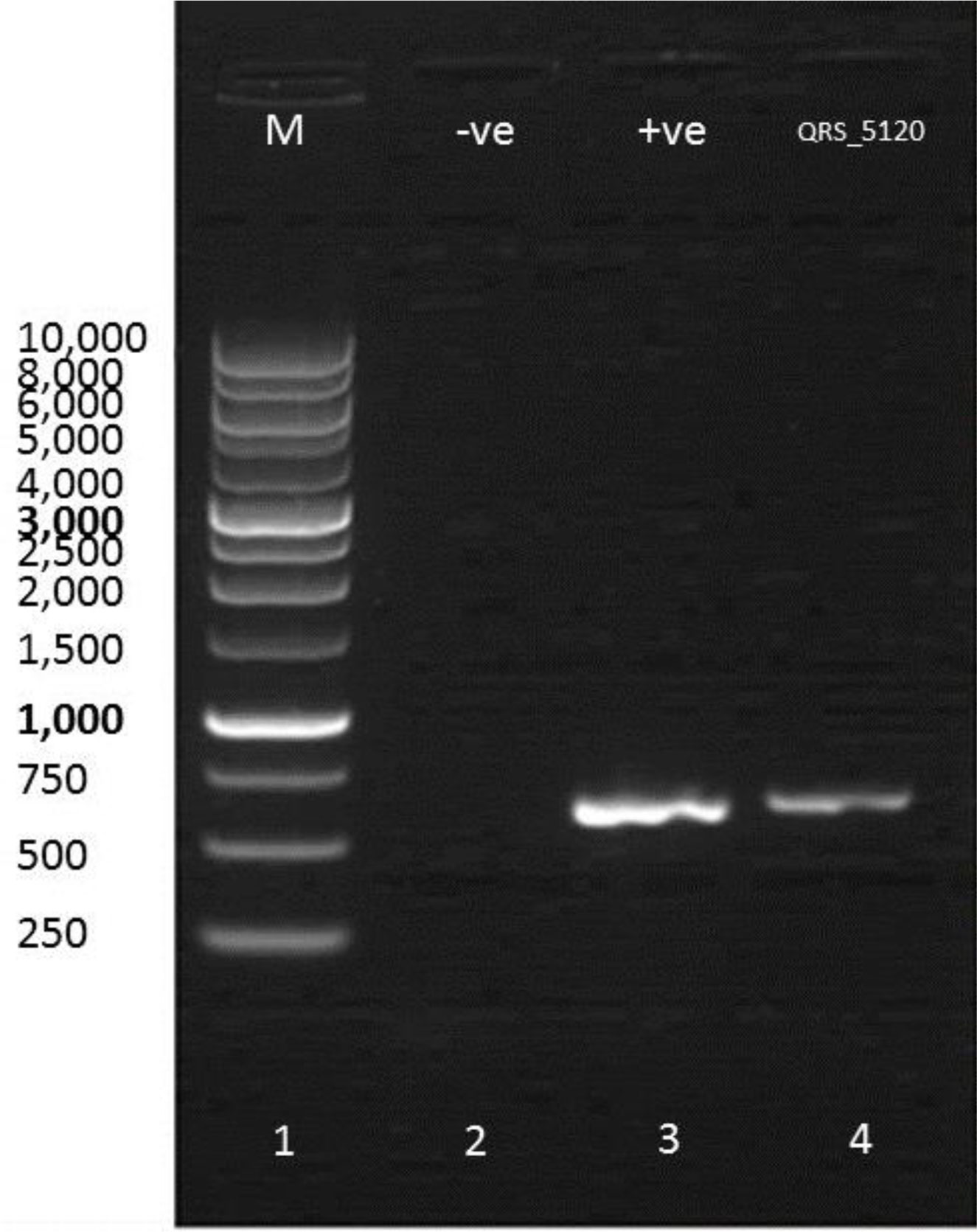
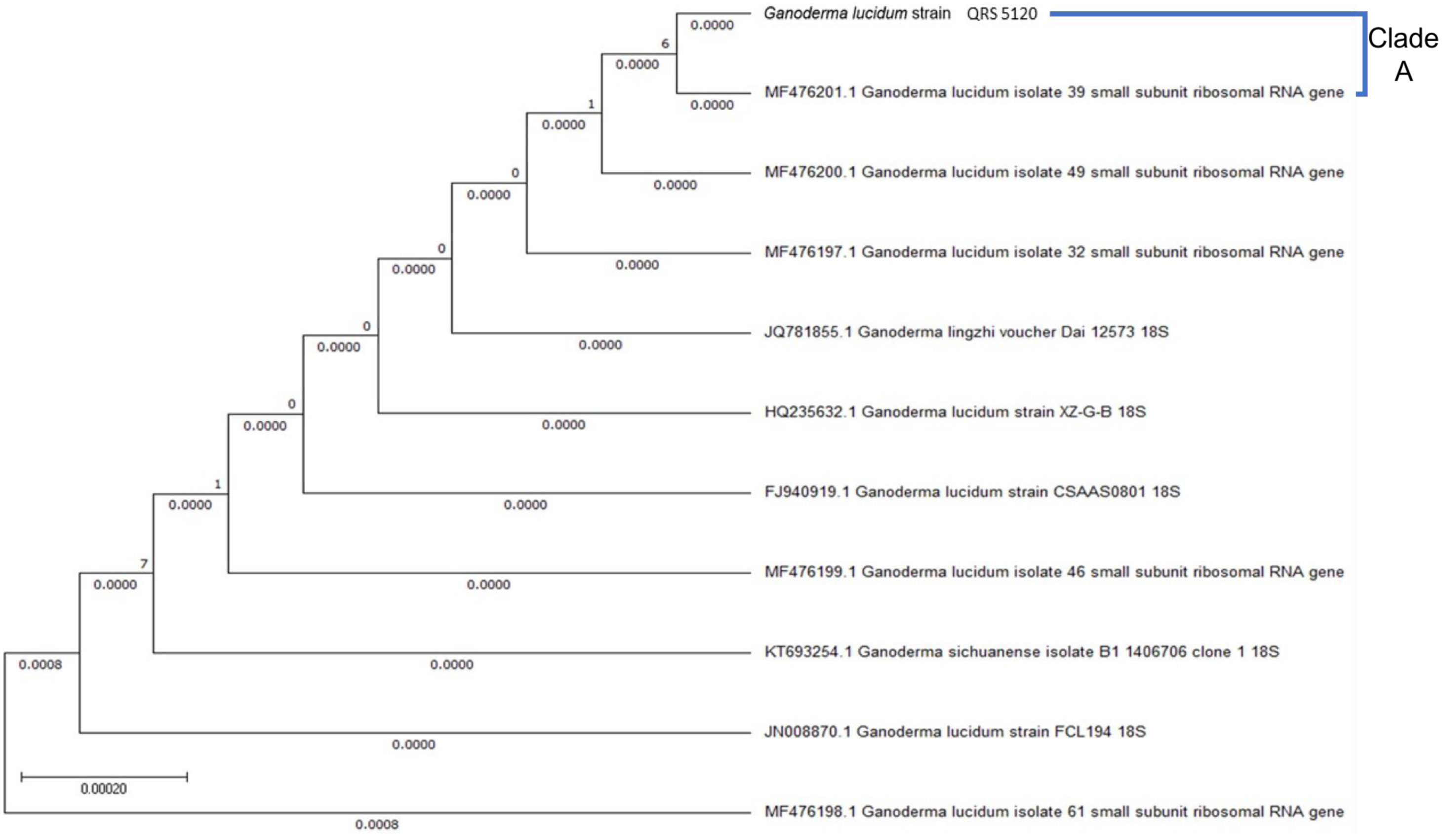
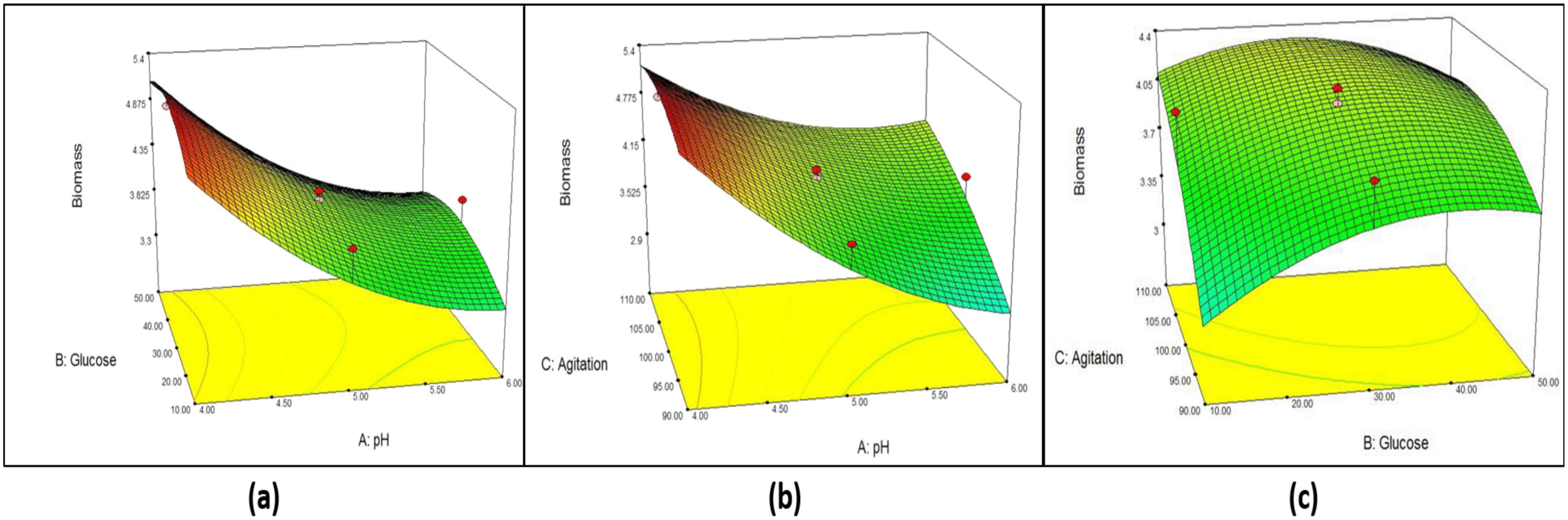
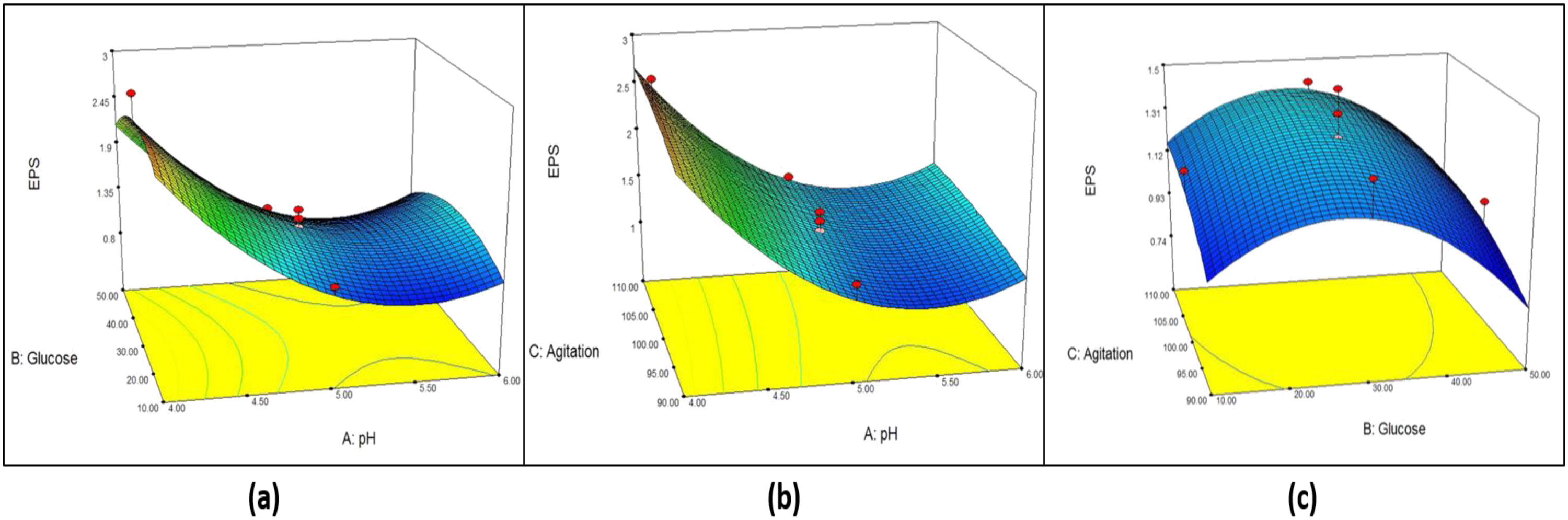
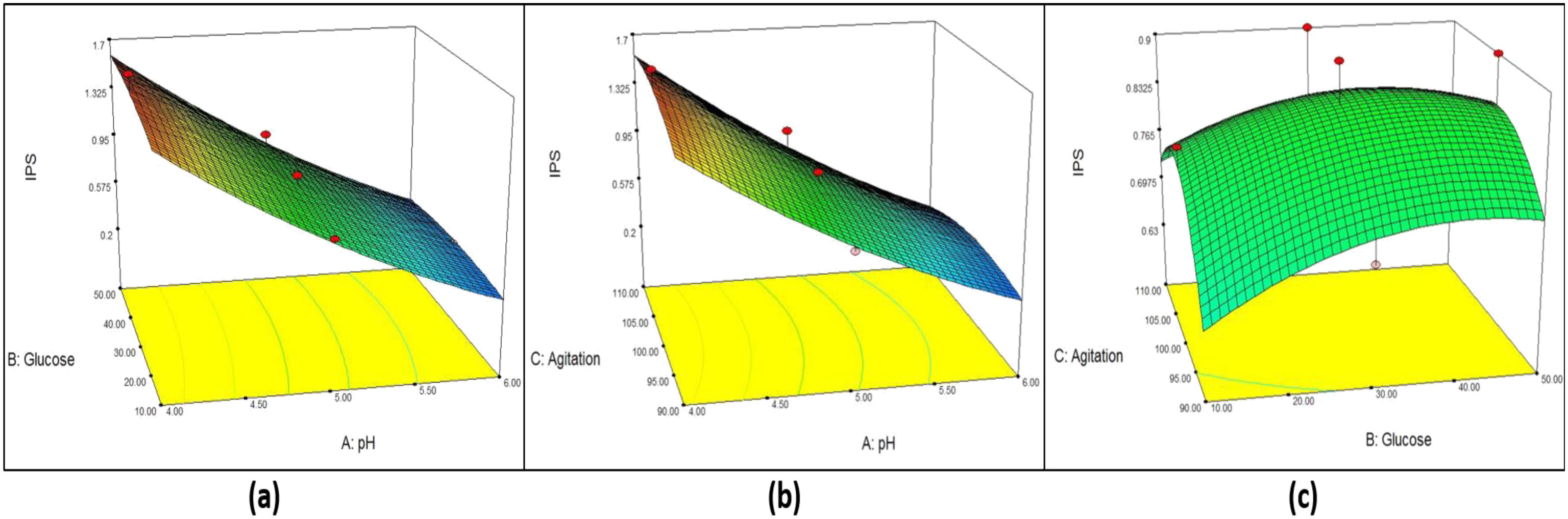
Figures(6) / Tables(7)

Sugenendran Supramani, Rahayu Ahmad, Zul Ilham, Mohamad Suffian Mohamad Annuar, Anita Klaus, Wan Abd Al Qadr Imad Wan-Mohtar. Optimisation of biomass, exopolysaccharide and intracellular polysaccharide production from the mycelium of an identified Ganoderma lucidum strain QRS 5120 using response surface methodology[J]. AIMS Microbiology, 2019, 5(1): 19-38. doi: 10.3934/microbiol.2019.1.19







 DownLoad:
DownLoad: