Citation: Luís Dias, Tânia Rosado, Ana Coelho, Pedro Barrulas, Luís Lopes, Patrícia Moita, António Candeias, José Mirão, Ana Teresa Caldeira. Natural limestone discolouration triggered by microbial activity—a contribution[J]. AIMS Microbiology, 2018, 4(4): 594-607. doi: 10.3934/microbiol.2018.4.594

| [1] |
Urosevic M, Sebastián E, Cardell C (2013) An experimental study on the influence of surface finishing on the weathering of a building low-porous limestone in coastal environments. Eng Geol 154: 131–141. doi: 10.1016/j.enggeo.2012.12.013

|
| [2] |
Mihajlovski A, Gabarre A, Seyer D, et al. (2017) Bacterial diversity on rock surface of the ruined part of a French historic monument: the Chaalis abbey. Int Biodeter Biodegr 120: 161–169. doi: 10.1016/j.ibiod.2017.02.019
|
| [3] |
Rosado T, Reis A, Mirão J, et al. (2014) Pink! Why not? On the unusual color of Évora Cathedral. Int Biodeter Biodegr 94: 121–127. doi: 10.1016/j.ibiod.2014.07.010
|
| [4] |
Rosado T, Gil M, Mirão J, et al. (2013) Oxalate biofilm formation in mural paintings due to microorganisms-A comprehensive study. Int Biodeter Biodegr 85: 1–7. doi: 10.1016/j.ibiod.2013.06.013
|
| [5] | Gaylarde P, Gaylarde C (2004) Deterioration of siliceous stone monuments in Latin America: Microorganisms and mechanisms. Corros Rev 22: 395–415. |
| [6] |
Gaylarde CC, Ortega-Morales BO, Bartolo-Perez P (2007) Biogenic black crusts on buildings in unpolluted environments. Curr Microbiol 54: 162–166. doi: 10.1007/s00284-006-0432-8
|
| [7] |
Scheerer S, Ortega-Morales O, Gaylarde C (2009) Microbial deterioration of stone monuments-An updated overview. Adv Appl Microbiol 66: 97–139. doi: 10.1016/S0065-2164(08)00805-8
|
| [8] |
Urzì C, De Leo F, Krakova L, et al. (2016) Effects of biocide treatments on the biofilm community in Domitilla's catacombs in Rome. Sci Total Environ 572: 252–262. doi: 10.1016/j.scitotenv.2016.07.195
|
| [9] | Li Q, Zhang B, He Z, et al. (2016) Distribution and diversity of bacteria and fungi colonization in stone monuments analysed by high-throughput sequencing. PLoS One 11: e0163287. |
| [10] |
Nowicka-Krawczyk P, Zelazna-Wieczorek J, Otlewska A, et al. (2014) Diversity of an aerial phototrophic coating of historic buildings in the former Auschwitz II-Birkenau concentration camp. Sci Total Environ 493: 116–123. doi: 10.1016/j.scitotenv.2014.05.113
|
| [11] |
Borderie F, Denis M, Barani A, et al. (2016) Microbial composition and ecological features of phototrophic biofilms proliferating in the Moidons Caves (France): investigation at the single-cell level. Environ Sci Pollut R 23: 12039–12049. doi: 10.1007/s11356-016-6414-x
|
| [12] |
Albertano P, Urzi C (1999) Structural interactions among epilithic cyanobacteria and heterotrophic microorganisms in Roman hypogea. Microb Ecol 38: 244–252. doi: 10.1007/s002489900174
|
| [13] |
Mosmann T (1983) Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J Immunol Methods 65: 55–63. doi: 10.1016/0022-1759(83)90303-4
|
| [14] |
Herlemann DP, Labrenz M, Jurgens K, et al. (2011) Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J 5: 1571–1579. doi: 10.1038/ismej.2011.41
|
| [15] |
Klindworth A, Pruesse E, Schweer T, et al. (2013) Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res 41: e1. doi: 10.1093/nar/gks808
|
| [16] | Tedersoo L, Bahram M, Polme S, et al. (2014) Global diversity and geography of soil fungi. Science 346: 1078. |
| [17] |
Schmieder R, Edwards R (2011) Quality control and preprocessing of metagenomic datasets. Bioinformatics 27: 863–864. doi: 10.1093/bioinformatics/btr026
|
| [18] |
Schubert M, Lindgreen S, Orlando L (2016) AdapterRemoval v2: rapid adapter trimming, identification, and read merging. BMC Res Notes 9: 88. doi: 10.1186/s13104-016-1900-2
|
| [19] |
Caporaso JG, Kuczynski J, Stombaugh J, et al. (2010) QIIME allows analysis of high-throughput community sequencing data. Nat Methods 7: 335–336. doi: 10.1038/nmeth.f.303
|
| [20] |
Edgar RC, Haas BJ, Clemente JC, et al. (2011) UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 27: 2194–2200. doi: 10.1093/bioinformatics/btr381
|
| [21] |
DeSantis TZ, Hugenholtz P, Larsen N, et al. (2006) Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl Environ Microbiol 72: 5069–5072. doi: 10.1128/AEM.03006-05
|
| [22] |
Abarenkov K, Henrik NR, Larsson K, et al. (2010) The UNITE database for molecular identification of fungi-recent updates and future perspectives. New Phytol 186: 281–285. doi: 10.1111/j.1469-8137.2009.03160.x
|
| [23] |
Muynck W, Leuridan S, Van Loo D, et al. (2011) Influence of Pore Structure on the effectiveness of a biogenic carbonate surface treatment for limestone conservation. Appl Environ Microbiol 77: 6808–6820. doi: 10.1128/AEM.00219-11
|
| [24] | Rosado T, Martins MM, Pires M, et al. (2013) Enzymatic monitorization of mural paintings biodegradation and biodeterioration. Int J Conserv Sci 4: 603–612. |
| [25] |
Vázquez-Nion D, Rodríguez-Castro J, López-Rodríguez MC, et al. (2016) Subaerial biofilms on granitic historic buildings: microbial diversity and development of phototrophic multi-species cultures. Biofouling 32: 657–669. doi: 10.1080/08927014.2016.1183121
|
| [26] |
Ogawa A, Celikkol-Aydin S, Gaylarde C, et al. (2017) Microbiomes of biofilms on decorative siliceous stone: drawbacks and advantages of next generation sequencing. Curr Microbiol 74: 848–853. doi: 10.1007/s00284-017-1257-3
|
| [27] | Ettenauer J, Jurado V, Piñar G, et al. (2014) Halophilic microorganisms are responsible for the rosy discolouration of saline environments in three historical buildings with mural paintings. PLoS One 9: e103844. |
| [28] |
Urzi C, Brusetti L, Salamone P, et al. (2001) Biodiversity of Geodermatophilaceae isolated from altered stones and monuments in the Mediterranean basin. Environ Microbiol 3: 471–479. doi: 10.1046/j.1462-2920.2001.00217.x
|
| [29] |
Gaylarde C, Baptista-Neto JA, Ogawa A, et al. (2017) Epilithic and endolithic microorganisms and deterioration on stone church facades subject to urban pollution in a sub-tropical climate. Biofouling 33: 113–127. doi: 10.1080/08927014.2016.1269893
|
| [30] |
Rosado T, Mirão J, Candeias A, et al. (2014) Microbial communities analysis assessed by pyrosequencing-a new approach applied to conservation state studies of mural paintings. Anal Bioanal Chem 406: 887–895. doi: 10.1007/s00216-013-7516-7
|


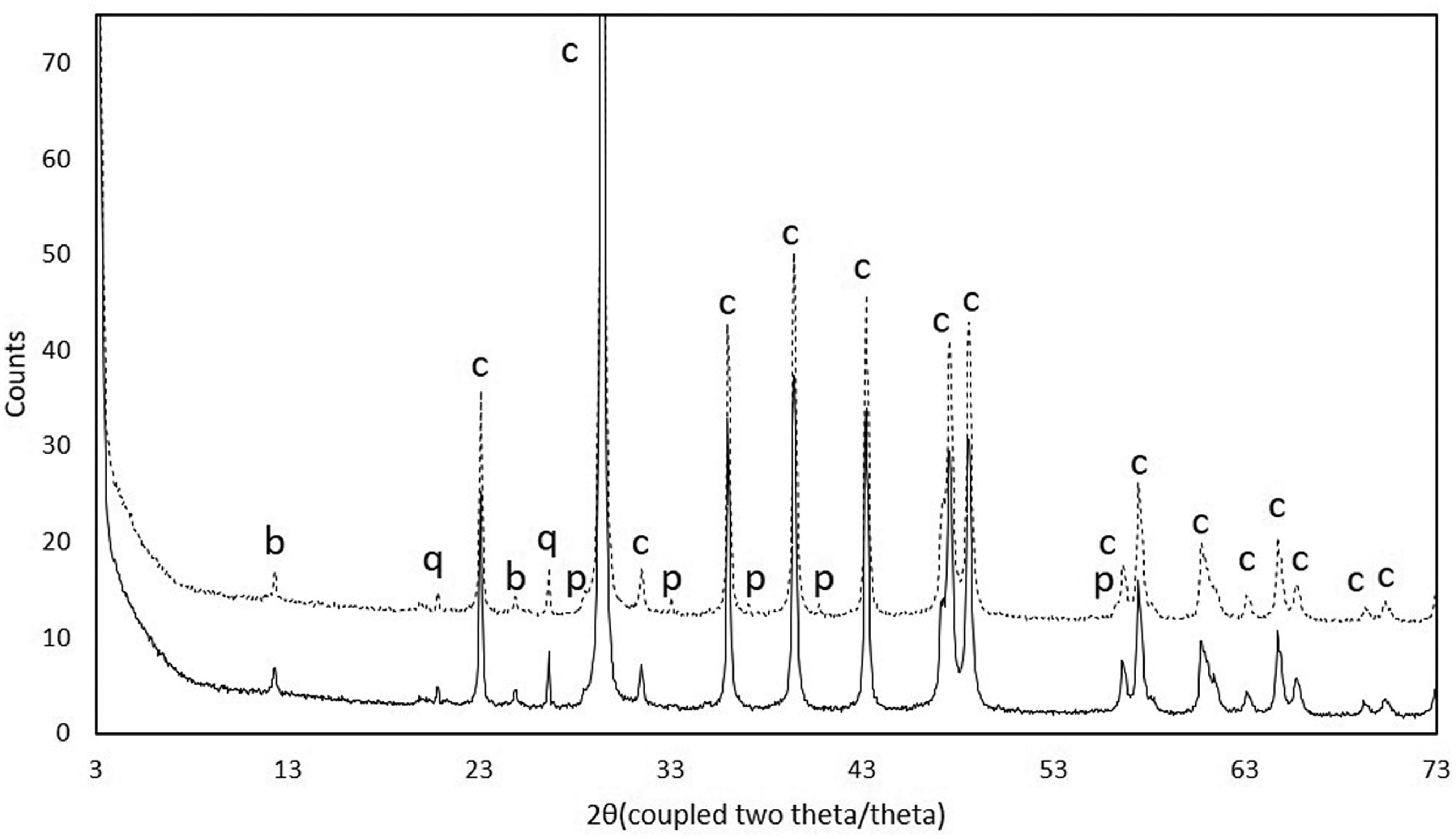
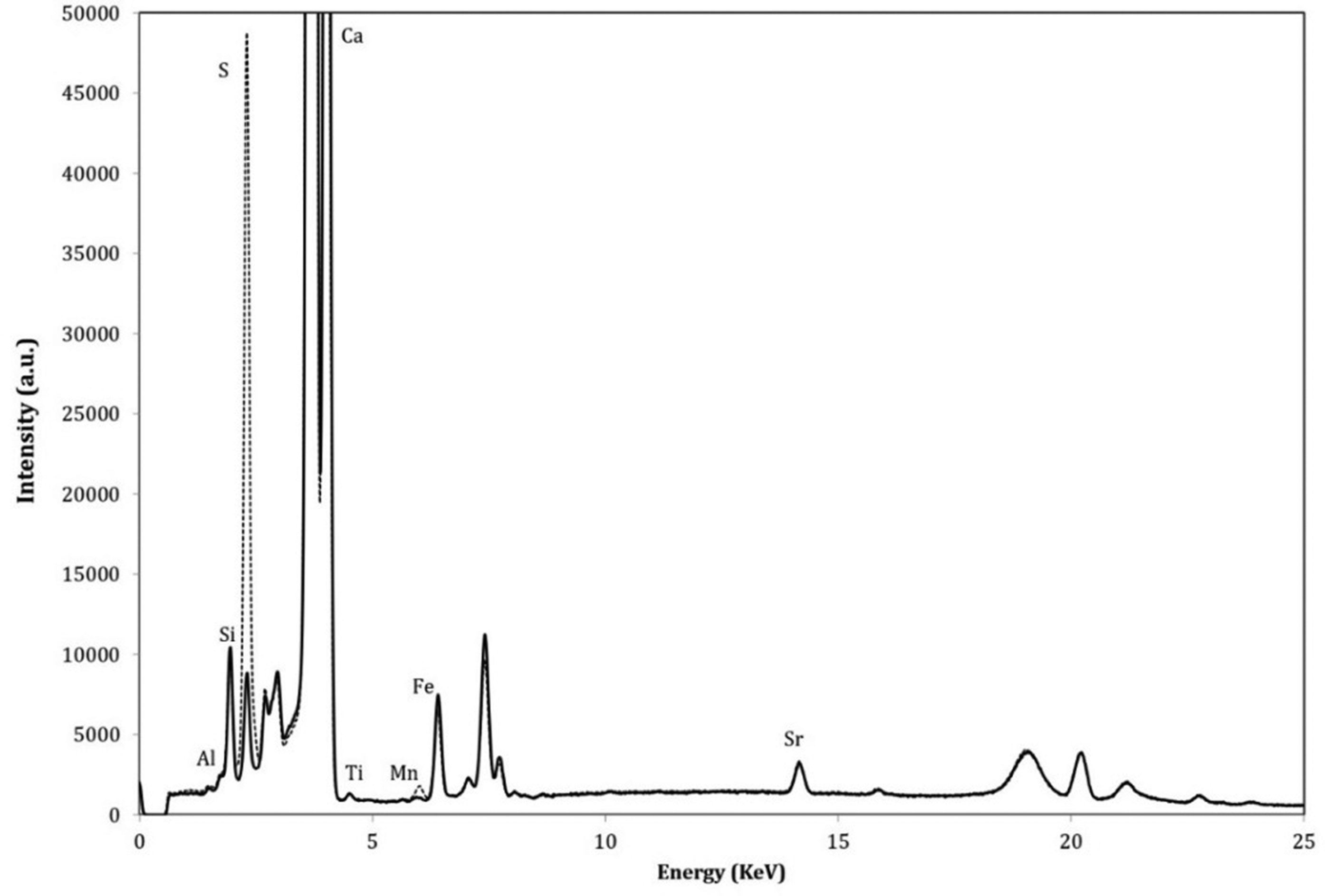
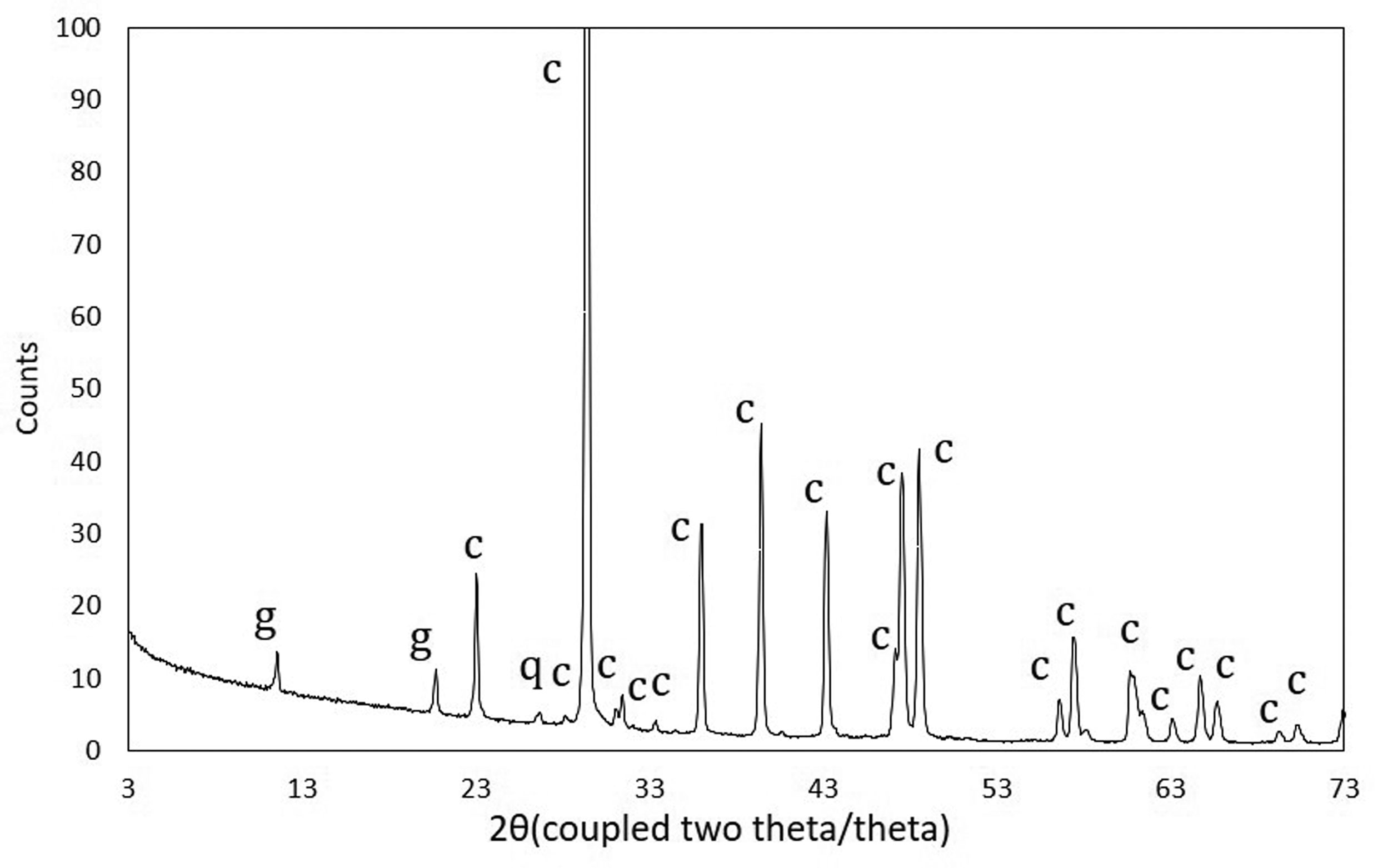
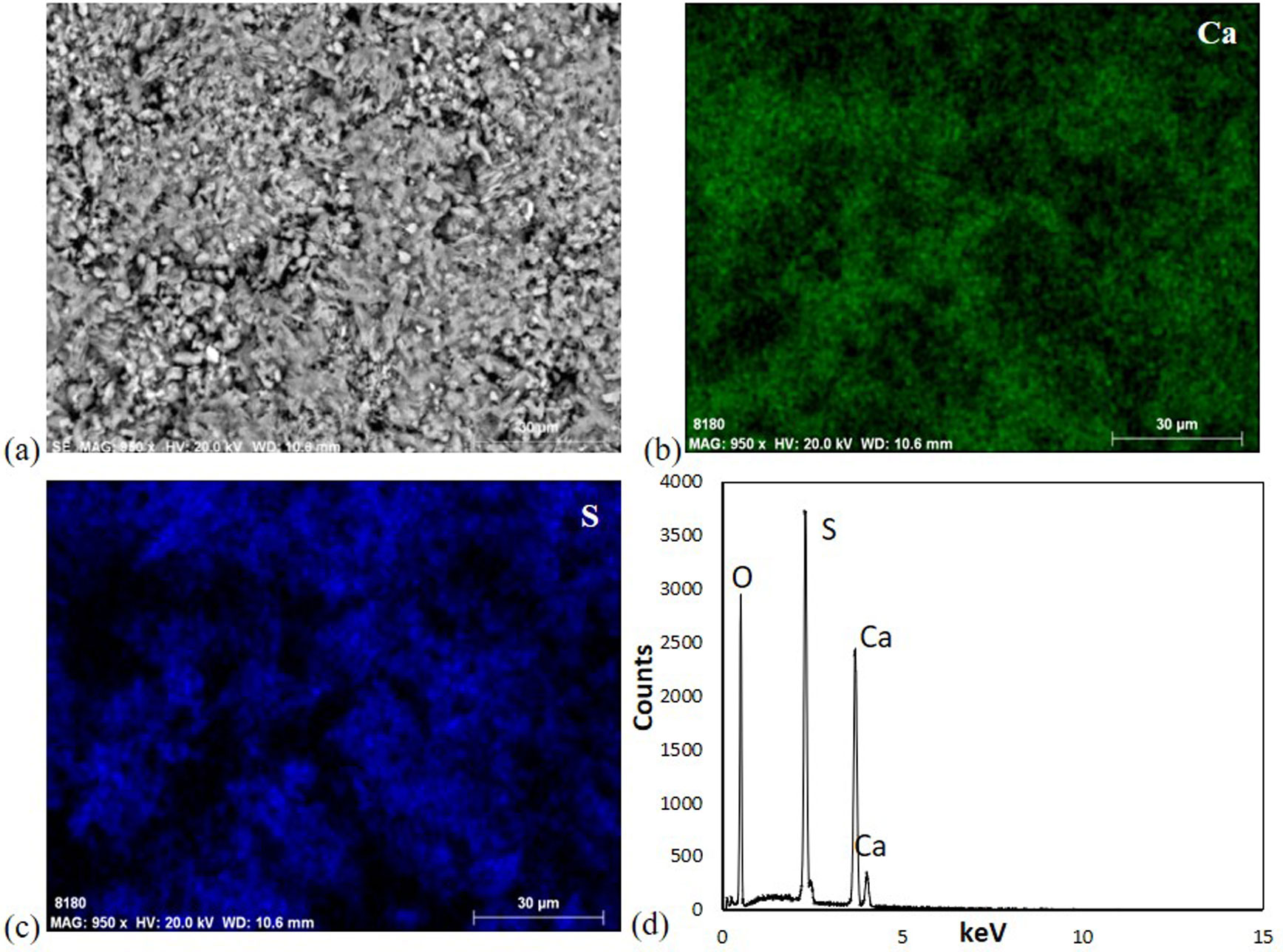


Figures(11)

Luís Dias, Tânia Rosado, Ana Coelho, Pedro Barrulas, Luís Lopes, Patrícia Moita, António Candeias, José Mirão, Ana Teresa Caldeira. Natural limestone discolouration triggered by microbial activity—a contribution[J]. AIMS Microbiology, 2018, 4(4): 594-607. doi: 10.3934/microbiol.2018.4.594







 DownLoad:
DownLoad: