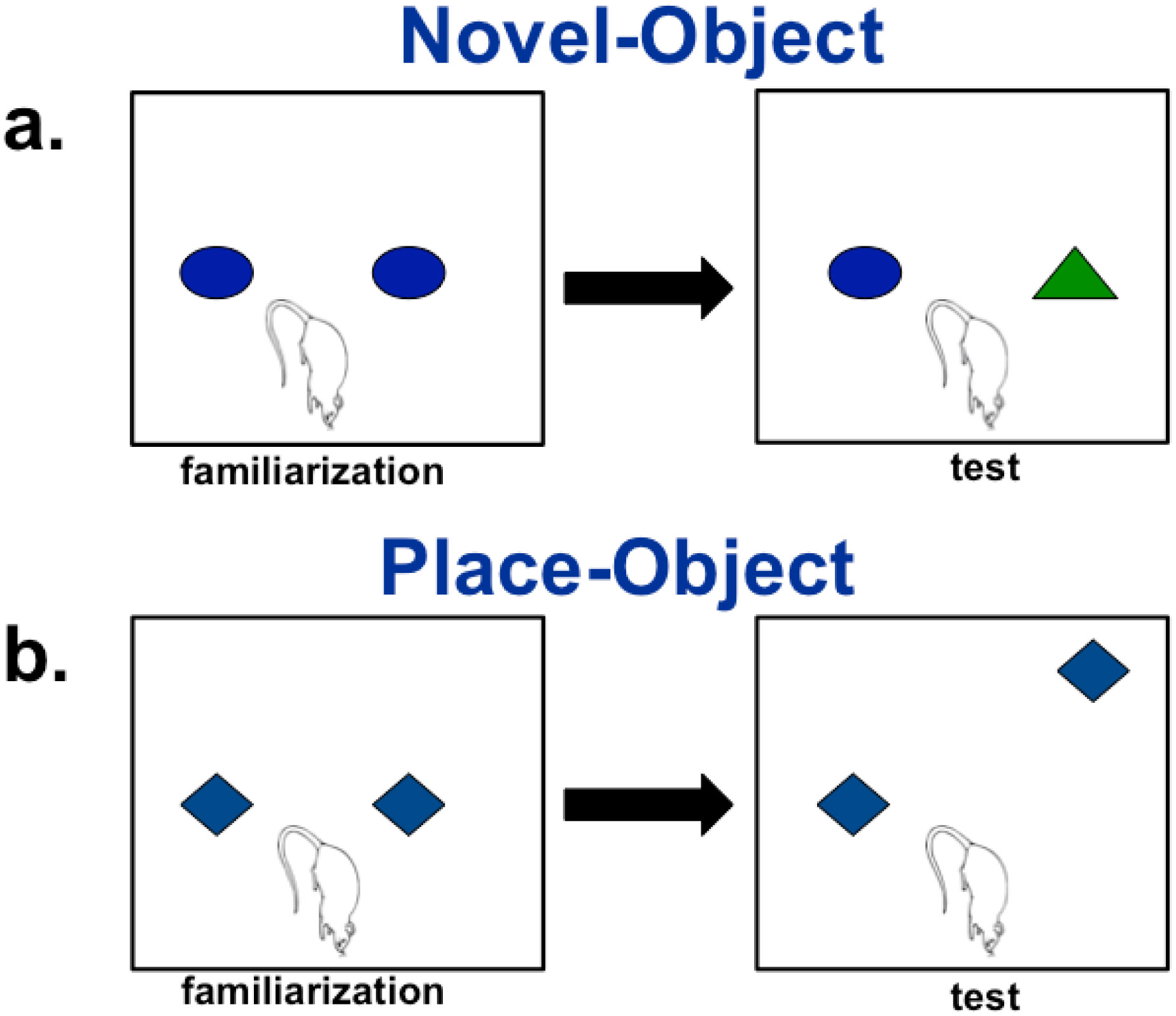
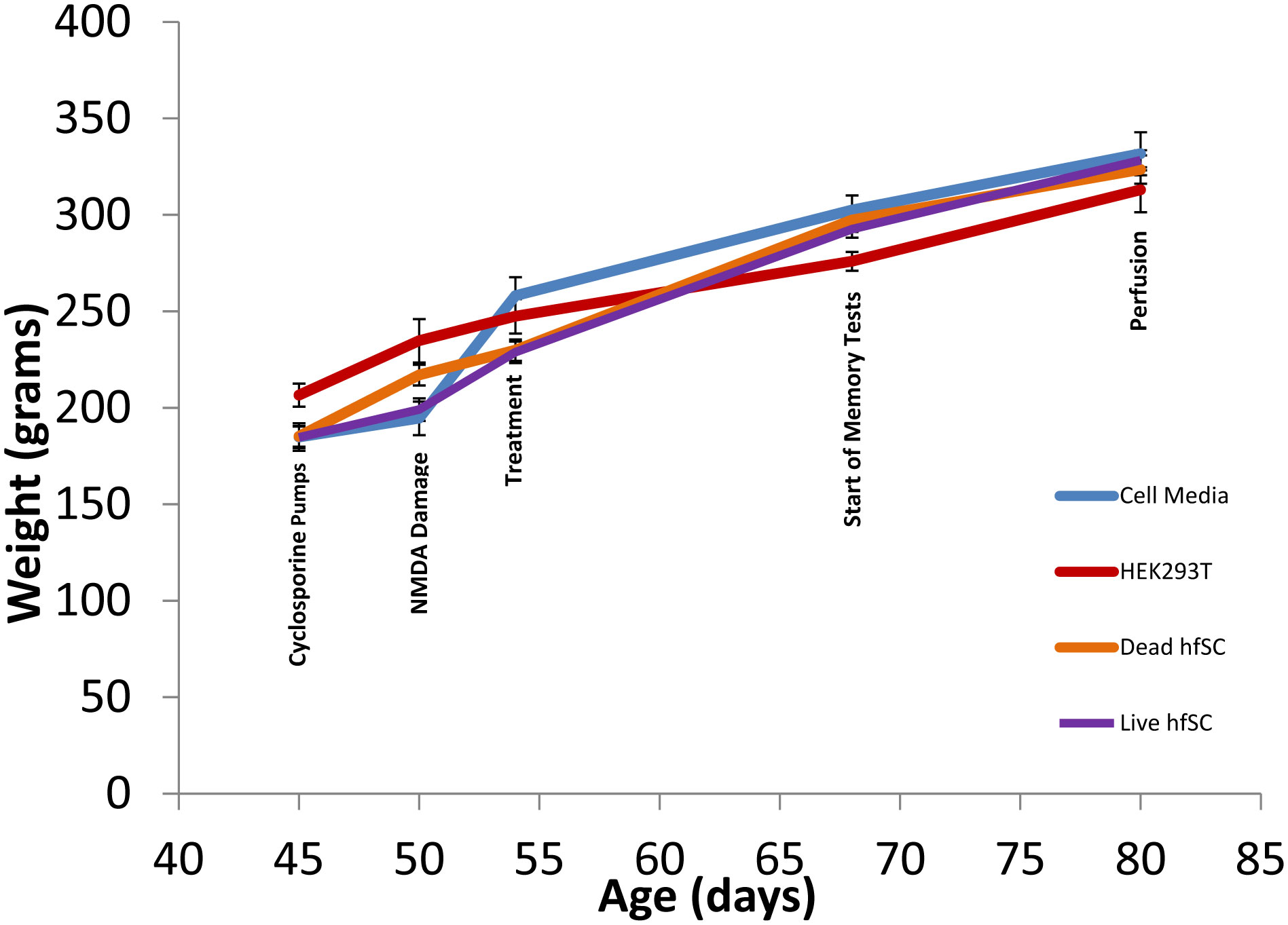
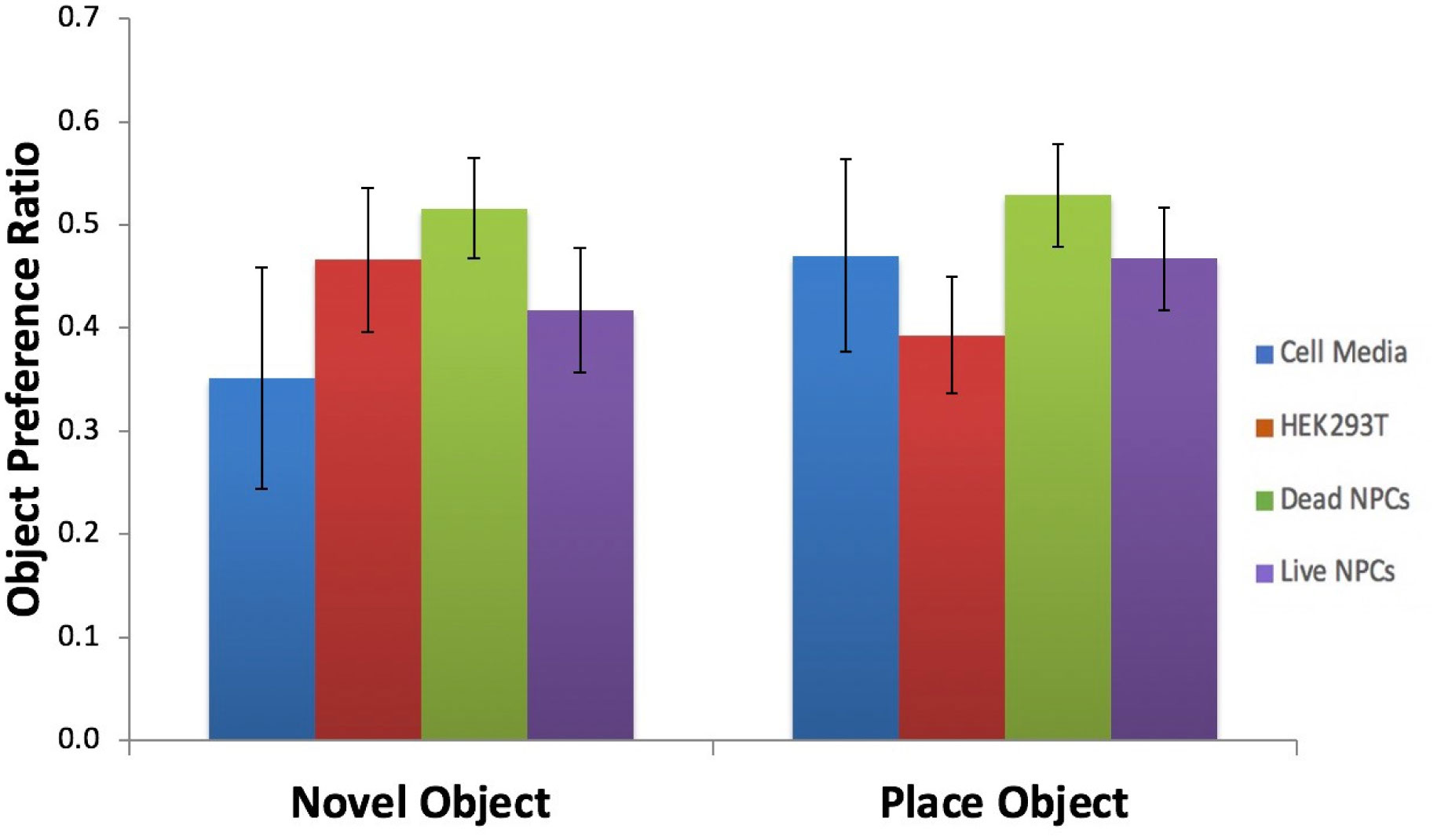
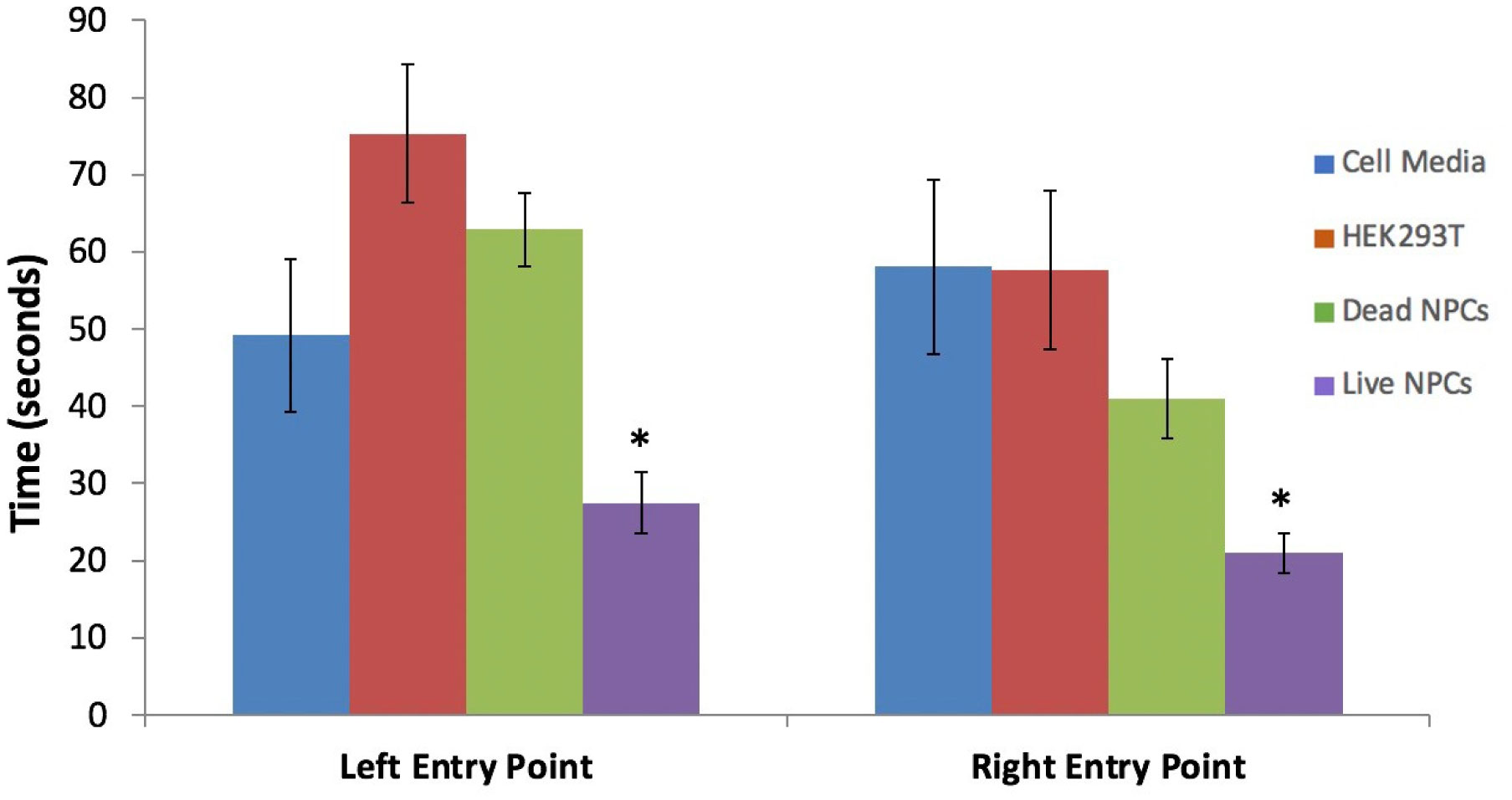
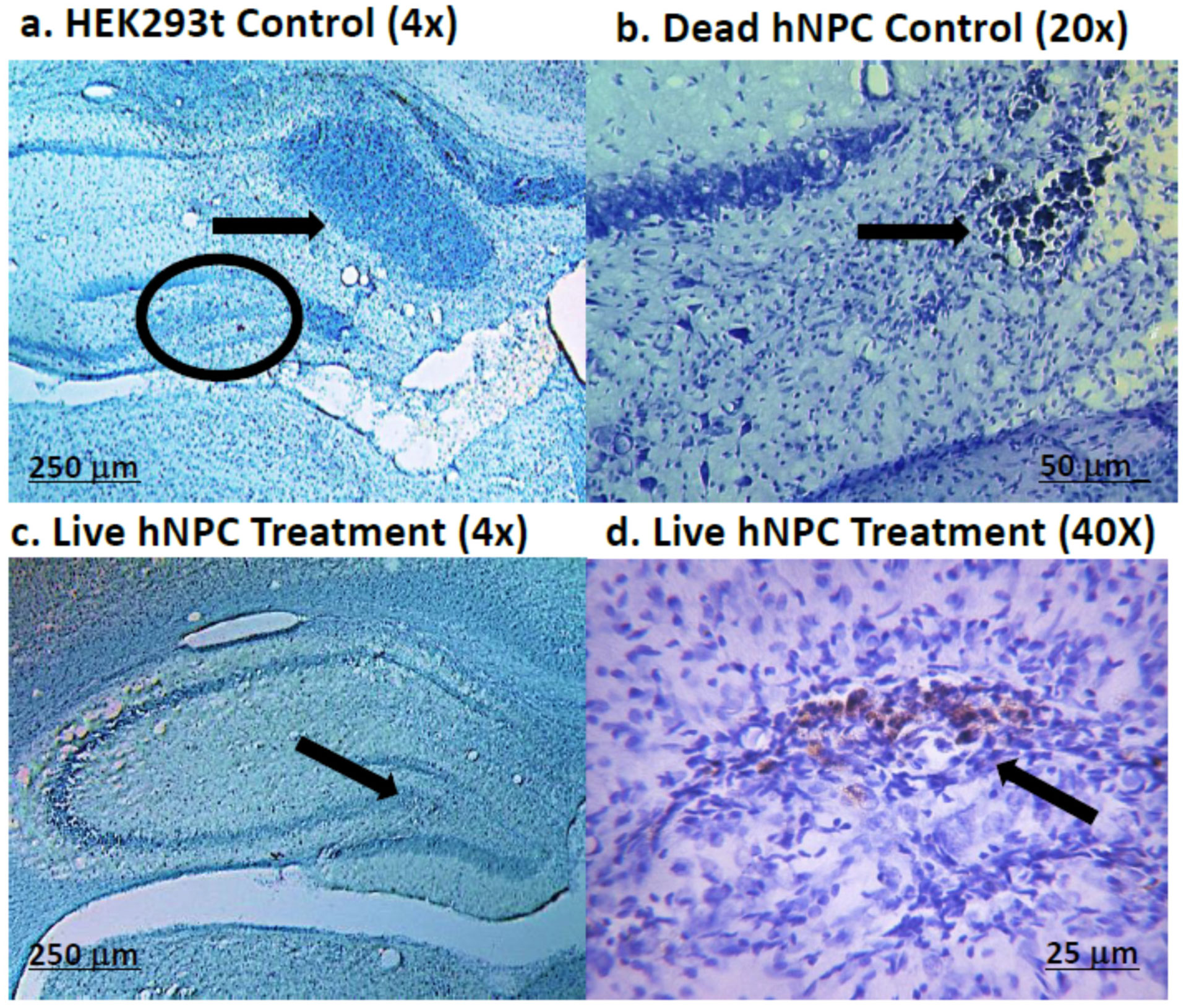
It has been established that the CA3 region of the hippocampus is involved in consolidating short-term memory to long-term memory and aids in spatial navigation retention. Seizures and many neurologic diseases induce damage to that region of the hippocampus, resulting in deficits in memory consolidation and spatial navigation. Drug treatments have been proven to have limited effectiveness, but cell replacement therapy has demonstrated to be more promising. Celavie Biosciences have developed a multipotent, nontumorigenic human neural progenitor cell (hNPC) line shown to have the ability to migrate in situ, reducing structural and functional deficits in neurodegenerative animal models. Here, we examined whether transplanted hNPCs would reestablish the memories of Han-Wistar rats subjected to hippocampal excitotoxic lesioning. The rats were lesioned in the CA3c regions at 50 days bilaterally with the neurotoxin NMDA (1 µl containing 7.5 mg/ml; −3.5 mm AP; ±2.0 L and −2.5 V). At 54 days of age, live hNPCs (500000 cells in 5 µl cell suspension media), frozen-killed hNPCs (500000 cells/5 µl), HEK293T cells (500000 cells/5 µl) or vehicle (cell suspension media; 5 µl) were bilaterally implanted directly into the NMDA damaged area. The rats were tested two weeks later with three different memory tests: novel and place-object assays and the water-maze task. Results showed that rats receiving live hNPC implantation performed significantly better in the water maze task than control groups; yet, novel and place-object test results showed no significant differences among treatments. Histology confirmed the survival of implanted hNPCs after 28 days post-implantation as well as showing neuroprotective effects. This study showed that Celavie's hNPCs were able to survive and improve some but not all hippocampal functionality, emphasizing the promise for cell replacement therapeutics for neurodegenerative disorders.
Citation: Sabrina K Uppal, Toni L Uhlendorf, Ruslan L Nuryyev, Jacqueline Saenz, Menaga Shanmugam, Jessica Ochoa, William Van Trigt, Cindy S Malone, Andrew P St. Julian, Oleg Kopyov, Alex Kopyov, Randy W Cohen. Human neural progenitor cells ameliorate NMDA-induced hippocampal degeneration and related functional deficits[J]. AIMS Medical Science, 2021, 8(3): 252-268. doi: 10.3934/medsci.2021021
It has been established that the CA3 region of the hippocampus is involved in consolidating short-term memory to long-term memory and aids in spatial navigation retention. Seizures and many neurologic diseases induce damage to that region of the hippocampus, resulting in deficits in memory consolidation and spatial navigation. Drug treatments have been proven to have limited effectiveness, but cell replacement therapy has demonstrated to be more promising. Celavie Biosciences have developed a multipotent, nontumorigenic human neural progenitor cell (hNPC) line shown to have the ability to migrate in situ, reducing structural and functional deficits in neurodegenerative animal models. Here, we examined whether transplanted hNPCs would reestablish the memories of Han-Wistar rats subjected to hippocampal excitotoxic lesioning. The rats were lesioned in the CA3c regions at 50 days bilaterally with the neurotoxin NMDA (1 µl containing 7.5 mg/ml; −3.5 mm AP; ±2.0 L and −2.5 V). At 54 days of age, live hNPCs (500000 cells in 5 µl cell suspension media), frozen-killed hNPCs (500000 cells/5 µl), HEK293T cells (500000 cells/5 µl) or vehicle (cell suspension media; 5 µl) were bilaterally implanted directly into the NMDA damaged area. The rats were tested two weeks later with three different memory tests: novel and place-object assays and the water-maze task. Results showed that rats receiving live hNPC implantation performed significantly better in the water maze task than control groups; yet, novel and place-object test results showed no significant differences among treatments. Histology confirmed the survival of implanted hNPCs after 28 days post-implantation as well as showing neuroprotective effects. This study showed that Celavie's hNPCs were able to survive and improve some but not all hippocampal functionality, emphasizing the promise for cell replacement therapeutics for neurodegenerative disorders.
Diaminobenzidine
Human neural progenitor cell
Insulin-like growth factor-1
Phosphate buffer saline
Paraformaldehyde
Subventricular zone-derived

| [1] |
Faber M, Mills C (2018) The critical role of the hippocampus in mind wandering. J Neurosci 38: 6439-6441. doi: 10.1523/JNEUROSCI.0995-18.2018

|
| [2] |
Anand KS, Dhikav V (2012) Hippocampus in health and disease: an overview. Ann Indian Acad Neurol 15: 239-246. doi: 10.4103/0972-2327.104323
|
| [3] |
Camina E, Güell F (2017) The neuroanatomical, neurophysiological and psychological basis of memory: current models and their origins. Front Pharmacol 8: 438. doi: 10.3389/fphar.2017.00438
|
| [4] |
Guzman SJ, Schlogl A, Frotscher M, et al. (2016) Synaptic mechanisms of pattern completion in the hippocampal CA3 network. Science 353: 1117-1123. doi: 10.1126/science.aaf1836
|
| [5] |
Maia GH, Quesado JL, Soares JI, et al. (2014) Loss of hippocampal neurons after kainate treatment correlates with behavioral deficits. PloS One 9: e84722. doi: 10.1371/journal.pone.0084722
|
| [6] |
Cherubini E, Miles R (2015) The CA3 region of the hippocampus: how is it? What is it for? How does it do it? Front Cell Neurosci 9: 19. doi: 10.3389/fncel.2015.00019
|
| [7] |
Hattiangady B, Shetty A (2012) Neural stem cell grafting counteracts hippocampal injury-mediated impairments in mood, memory and neurogenesis. Stem Cells Transl Med 1: 696-708. doi: 10.5966/sctm.2012-0050
|
| [8] |
Hattiangady B, Rao MS, Shetty AK (2004) Chronic temporal lobe epilepsy is associated with severely declined dentate neurogenesis in the adult hippocampus. Neurobiol Dis 17: 473-490. doi: 10.1016/j.nbd.2004.08.008
|
| [9] | Waldau B, Hattiangady B, Kuruba R, et al. (2010) Medial ganglionic eminence-derived neural stem cell grafts ease spontaneous seizures and restore GDNF expression in a rat model of chronic temporal lobe epilepsy. Stem Cells 28: 1153-1164. |
| [10] |
Park D, Joo SS, Kim TK, et al. (2012) Human neural stem cells overexpressing choline acetyltransferase restore cognitive function of kainic acid-induced learning and memory deficit animals. Cell Transplant 21: 365-371. doi: 10.3727/096368911X586765
|
| [11] |
Miltiadous P, Kouroupi G, Stamatakis A, et al. (2013) Subventricular zone-derived neural stem cell grafts protect against hippocampal degeneration and restore cognitive function in the mouse following intrahippocampal kainic acid administration. Stem Cells Transl Med 2: 185-198. doi: 10.5966/sctm.2012-0074
|
| [12] |
Kopyov A, Uhlendorf T, Cohen RW (2021) Why use pre-differentiated cells to address complex multi-factorial neurodegenerative diseases? Neural Regen Res 16: 1413-1414. doi: 10.4103/1673-5374.300990
|
| [13] |
Uhlendorf T, Nuryyev R, Kopyov A, et al. (2017) Efficacy of two delivery routes for transplanting human neural progenitor cells (NPCs) into the spastic Han-Wistar rat, a model of ataxia. Cell Transplant 26: 259-269. doi: 10.3727/096368916X693527
|
| [14] |
Nuryyev R, Uhlendorf T, Tierney W, et al. (2017) Transplantation of human neural progenitor cells reveals structural and functional improvements in the spastic Han-Wistar rat model of ataxia. Cell Transplant 26: 1811-1821. doi: 10.1177/0963689717723637
|
| [15] |
Tierney W, Uhlendorf T, Lemus A, et al. (2020) Transplanted human neural progenitor cells attenuate motor dysfunction and lengthen longevity in a rat model of ataxia. Cell Transplant 29: 1-12. doi: 10.1177/0963689720920275
|
| [16] |
Shen C, Gu M, Song C, et al. (2008) The tumorigenicity diversification in human embryonic kidney 293 cell line cultured in vitro. Biologicals 36: 263-268. doi: 10.1016/j.biologicals.2008.02.002
|
| [17] | Gilbert PE, Brushfield AM (2009) The role of the CA3 hippocampal subregion in spatial memory: a process oriented behavior assessment. Prog Neuropsychopharm Bio Psych 169: 142-149. |
| [18] |
Florian C, Roullet P (2004) Hippocampal CA3-region is crucial for acquisition and memory consolidation in Morris water maze task in mice. Behav Brain Res 154: 365-374. doi: 10.1016/j.bbr.2004.03.003
|
| [19] |
Ceccom J, Bouhsira E, Halley H, et al. (2013) Differential needs of zinc in the CA3 area of dorsal hippocampus for the consolidation of contextual fear and spatial memories. Learn Mem 20: 348-351. doi: 10.1101/lm.029017.112
|
| [20] |
Kelly S, Bliss TM, Shah AK, et al. (2004) Transplanted human fetal neural stem cells survive, migrate, and differentiate in ischemic rat cerebral cortex. Proc Natl Acad Sci USA 101: 11839-11844. doi: 10.1073/pnas.0404474101
|
| [21] |
Chang DJ, Oh SH, Lee N, et al. (2013) Contralaterally transplanted human embryonic stem cell-derived neural precursor cells (ENStem-A) migrate and improve brain functions in stroke-damaged rats. Exp Mol Med 15: e53-e53. doi: 10.1038/emm.2013.93
|
| [22] |
Hurelbrink CB, Armstrong RJE, Dunnett SB, et al. (2002) Neural cells from primary human striatal xenografts migrate extensively in the adult rat CNS. European J Neurosci 15: 1255-1266. doi: 10.1046/j.1460-9568.2002.01959.x
|
| [23] |
Li H, Matsumoto K, Watanabe H (1999) Different effects of unilateral and bilateral hippocampal lesions in rats on the performance of radial maze and odor-paired associate tasks. Brain Research Bulletin 48: 113-119. doi: 10.1016/S0361-9230(98)00157-9
|
| [24] |
Zou LB, Yamada K, Sasa M, et al. (1999) Two phases of behavioral plasticity in rats following unilateral excitotoxic lesion of the hippocampus. Neuroscience 92: 819-826. doi: 10.1016/S0306-4522(99)00029-9
|
| [25] |
Cavazos JE, Jones SM, Cross DJ (2004) Sprouting and synaptic reorganization in the subiculum and CA1 region of the hippocampus in acute and chronic models of partial-onset epilepsy. Neuroscience 126: 677-688. doi: 10.1016/j.neuroscience.2004.04.014
|
| [26] |
Cheng M (2013) Hypothalamic neurogenesis in the adult brain. Front Neuroendocrin 34: 167-178. doi: 10.1016/j.yfrne.2013.05.001
|
| [27] |
Jorge RE, Acio L, Starkstein SE, et al. (2007) Hippocampal volume and mood disorders after traumatic brain injury. Biol Psychiatry 62: 332-338. doi: 10.1016/j.biopsych.2006.07.024
|
| [28] |
Potvin O, Allen K, Thibaudeau G, et al. (2006) Performance on spatial working memory tasks after dorsal or ventral hippocampal lesions and adjacent damage to the subiculum. Behav Neurosci 120: 413-422. doi: 10.1037/0735-7044.120.2.413
|
| [29] |
Sun D, McGinn MJ, Zhou S, et al. (2007) Anatomical integration of newly generated dentate granule neurons following traumatic brain injury in adult rats and its association to cognitive recovery. Exp Neurol 204: 264-272. doi: 10.1016/j.expneurol.2006.11.005
|
| [30] |
Hester MS, Danzer SC (2013) Accumulation of abnormal adult-generated hippocampal granule cells predicts seizure frequency and severity. J Neurosci 33: 8926-8936. doi: 10.1523/JNEUROSCI.5161-12.2013
|
| [31] |
Shetty AK (2014) Hippocampal injury-induced cognitive and mood dysfunction, altered neurogenesis, and epilepsy: Can early neural stem cell grafting intervention provide protection? Epilepsy Behav 38: 117-124. doi: 10.1016/j.yebeh.2013.12.001
|
| [32] |
Boldrini M, Fulmore CA, Tartt AN, et al. (2018) Human hippocampal neurogenesis persists throughout aging. Cell Stem Cell 22: 589-599. doi: 10.1016/j.stem.2018.03.015
|
| [33] |
Aigner L, Arber S, Kapfhammer JP, et al. (1995) Overexpression of the neural growth-associated protein GAP-43 induces nerve sprouting in the adult nervous system of transgenic mice. Cell 83: 269-278. doi: 10.1016/0092-8674(95)90168-X
|
| [34] |
He Q, Dent EW, Meiri KF (1997) Modulation of actin filament behavior by GAP-43 (neuromodulin) is dependent on the phosphorylation status of serine 41, the protein kinase C site. J Neurosci 17: 3515-3524. doi: 10.1523/JNEUROSCI.17-10-03515.1997
|
| [35] |
Hughes-Davis EJ, Cogen JP, Jakowec MW, et al. (2005) Differential regulation of the growth-associated proteins GAP-43 and superior cervical ganglion 10 in response to lesions of the cortex and substantia nigra in the adult rat. Neuroscience 135: 1231-1239. doi: 10.1016/j.neuroscience.2005.07.017
|
| [36] |
Eskici NF, Erdem-Ozdamar S, Dayangac-Erden D (2018) The altered expression of perineuronal net elements during neural differentiation. Cell Mol Biol Lett 23: 1-12. doi: 10.1186/s11658-018-0073-5
|
| [37] |
Romberg C, Yang S, Melani R, et al. (2013) Depletion of perineuronal nets enhances recognition memory and long-term depression in the perirhinal cortex. J Neurosci 33: 7057-7065. doi: 10.1523/JNEUROSCI.6267-11.2013
|
| [38] |
Suttkus A, Rohn S, Weigel S, et al. (2014) Aggrecan, link protein and tenascin-R are essential components of the perineuronal net to protect neurons against iron-induced oxidative stress. Cell Death Dis 5: e1119. doi: 10.1038/cddis.2014.25
|
| [39] |
Wu JS, Cheung WM, Tsai YS, et al. (2009) Ligand-activated peroxisome proliferator-activated receptor-gamma protects against ischemic cerebral infarction and neuronal apoptosis by 14-3-3 epsilon upregulation. Circulation 119: 1124-1134. doi: 10.1161/CIRCULATIONAHA.108.812537
|
| [40] |
Yacoubian TA, Slone SR, Harrington AJ, et al. (2010) Differential neuroprotective effects of 14-3-3 proteins in models of Parkinson's disease. Cell Death Dis 1: e2. doi: 10.1038/cddis.2009.4
|
| [41] | Kristinsson J, Snaedal J, Tórsdóttir G, et al. (2012) Ceruloplasmin and iron in Alzheimer's disease and Parkinson's disease: a synopsis of recent studies. Neuropsychiatr Dis Treat 8: 515-521. |
| [42] | Montes S, Rivera-Mancia S, Diaz-Ruiz A, et al. (2014) Copper and copper proteins in Parkinson's disease. Oxid Med Cell Longev . |
| [43] |
Martín-Montañez E, Millon C, Boraldi F, et al. (2017) IGF-II promotes neuroprotection and neuroplasticity recovery in a long-lasting model of oxidative damage induced by glucocorticoids. Redox Biol 13: 69-81. doi: 10.1016/j.redox.2017.05.012
|
Figures(5)

Sabrina K Uppal, Toni L Uhlendorf, Ruslan L Nuryyev, Jacqueline Saenz, Menaga Shanmugam, Jessica Ochoa, William Van Trigt, Cindy S Malone, Andrew P St. Julian, Oleg Kopyov, Alex Kopyov, Randy W Cohen. Human neural progenitor cells ameliorate NMDA-induced hippocampal degeneration and related functional deficits[J]. AIMS Medical Science, 2021, 8(3): 252-268. doi: 10.3934/medsci.2021021







 DownLoad:
DownLoad: