Citation: Michiro Muraki. Sensitization to cell death induced by soluble Fas ligand and agonistic antibodies with exogenous agents: A review[J]. AIMS Medical Science, 2020, 7(3): 122-203. doi: 10.3934/medsci.2020011

| [1] |
Walczak H (2013) Death receptor-ligand systems in cancer, cell death, and inflammation. Cold Spring Harb Perspect Biol 5: a008698. doi: 10.1101/cshperspect.a008698

|
| [2] |
Calmon-Hamaty F, Audo R, Combe B, et al. (2015) Targeting the Fas/FasL system in rheumatoid arthritis therapy: promising or risky? Cytokine 75: 228-233. doi: 10.1016/j.cyto.2014.10.004
|
| [3] |
Franke DDH, Yolcu ES, Alard P, et al. (2007) A novel multimeric form of FasL modulates the ability of diabetogenic T cells to mediate type 1 diabetes in an adoptive transfer model. Mol Immunol 44: 2884-2892. doi: 10.1016/j.molimm.2007.01.014
|
| [4] |
Nagata S (1997) Apoptosis by death factor. Cell 88: 355-365. doi: 10.1016/S0092-8674(00)81874-7
|
| [5] |
Muraki M (2018) Development of expression systems for the production of recombinant human Fas ligand extracellular domain derivatives using Pichia pastoris and preparation of the conjugates by site-specific chemical modifications: A review. AIMS Bioengineer 5: 39-62. doi: 10.3934/bioeng.2018.1.39
|
| [6] |
Powell WC, Fingleton B, Wilson CL, et al. (1999) The metalloproteinase matrilysin proteolytically generates active soluble Fas ligand and potentiates epithelial cell apoptosis. Curr Biol 9: 1441-1447. doi: 10.1016/S0960-9822(00)80113-X
|
| [7] | Mitsiades N, Yu WH, Poulaki V, et al. (2001) Matrix metalloproteinase-7-mediated cleavage of Fas ligand protects tumor cells from chemotherapeutic drug cytotoxicity. Cancer Res 61: 577-581. |
| [8] |
Schneider P, Holler N, Bodmer JL, et al. (1998) Conversion of membrane-bound Fas (CD95) ligand to its soluble form is associated with downregulation of its proapoptotic activity and loss of liver toxicity. J Exp Med 187: 1205-1213. doi: 10.1084/jem.187.8.1205
|
| [9] |
Tanaka M, Suda T, Takahashi T, et al. (1995) Expression of functional soluble form of human Fas ligand in activated lymphocytes. EMBO J 14: 1129-1135. doi: 10.1002/j.1460-2075.1995.tb07096.x
|
| [10] |
Herrero R, Kajikawa O, Matute-Bello G, et al. (2011) The biological activity of FasL in human and mouse lungs is determined by the structure of its stalk region. J Clin Invest 121: 1174-1190. doi: 10.1172/JCI43004
|
| [11] |
Tanaka M, Itai T, Adachi M, et al. (1998) Downregulation of Fas ligand by shedding. Nature Med 4: 31-36. doi: 10.1038/nm0198-031
|
| [12] |
Matsumoto H, Murakami Y, Kataoka K, et al. (2015) Membrane-bound and soluble Fas ligands have opposite functions in photoreceptor cell death following separation from the retinal pigment epithelium. Cell Death Dis 6: e1986. doi: 10.1038/cddis.2015.334
|
| [13] |
Suda T, Hashimoto H, Tanaka M, et al. (1997) Membrane Fas ligand kills human peripheral blood T lymphocytes, and soluble Fas ligand blocks the killing. J Exp Med 186: 2045-2050. doi: 10.1084/jem.186.12.2045
|
| [14] |
Audo R, Calmon-Hamaty F, Papon L, et al. (2014) Distinct effects of soluble and membrane-bound Fas ligand on fibroblast-like synoviocytes from rheumatoid arthritis patients. Arthritis Rheum 66: 3289-3299. doi: 10.1002/art.38806
|
| [15] |
Song E, Chen J, Ouyang N, et al. (2001) Soluble Fas ligand released by colon adenocarcinoma cells induces host lymphocyte apoptosis: an active mode of immune evasion in colon cancer. Br J Cancer 85: 1047-1054. doi: 10.1054/bjoc.2001.2042
|
| [16] |
Wadsworth SJ, Atsuta R, McIntyre JO, et al. (2010) IL-13 and TH2 cytokine exposure triggers matrix metalloproteinase 7-mediated Fas ligand cleavage from bronchial epithelial cells. J Allergy Clin Immnunol 126: 366-374. doi: 10.1016/j.jaci.2010.05.015
|
| [17] |
Soni H, Kaminski D, Gangaraju R, et al. (2018) Cisplatin-induced oxidative stress stimulates renal Fas ligand shedding. Renal Failure 40: 314-322. doi: 10.1080/0886022X.2018.1456938
|
| [18] |
Lotti R, Shu E, Petrachi T, et al. (2018) Soluble Fas ligand is essential for blister formation in pemphigus. Front Immunol 9: 370. doi: 10.3389/fimmu.2018.00370
|
| [19] |
Komada Y, Inaba H, Li QS, et al. (1999) Epitopes and functional responses defined by a panel of anti-Fas (CD95) monoclonal antibodies. Hybridoma 18: 391-398. doi: 10.1089/hyb.1999.18.391
|
| [20] |
Chodorge M, Züger S, Stirnimann C, et al. (2012) A series of Fas receptor agonist antibodies that demonstrate an inverse correlation between affinity and potency. Cell Death Differ 19: 1187-1195. doi: 10.1038/cdd.2011.208
|
| [21] |
Shiraishi T, Suzuyama K, Okamoto H, et al. (2004) Increased cytotoxicity of soluble Fas ligand by fusing isoleucine zipper motif. Biochem Biophys Res Comm 322: 197-202. doi: 10.1016/j.bbrc.2004.07.098
|
| [22] |
Daburon S, Devaud C, Costet P, et al. (2013) Functional characterization of a chimeric soluble Fas ligand polymer with in vivo anti-tumor activity. Plos One 8: e54000. doi: 10.1371/journal.pone.0054000
|
| [23] |
Cremesti A, Paris F, Grassme H, et al. (2001) Ceramide enables Fas to cap and kill. J Biol Chem 276: 23954-23961. doi: 10.1074/jbc.M101866200
|
| [24] |
Muraki M (2014) Improved production of recombinant human Fas ligand extracellular domain in Pichia pastoris: yield enhancement using disposable culture-bag and its application to site-specific chemical modifications. BMC Biotechnol 14: 19. doi: 10.1186/1472-6750-14-19
|
| [25] |
Wajant H, Gerspach J, Pfizenmaier K (2013) Engineering death receptor ligands for cancer therapy. Cancer Lett 332: 163-174. doi: 10.1016/j.canlet.2010.12.019
|
| [26] |
Muraki M, Hirota K (2017) Site-specific chemical conjugation of human Fas ligand extracellular domain using trans-cyclooctene–methyltetrazine reactions. BMC Biotechnol 17: 56. doi: 10.1186/s12896-017-0381-2
|
| [27] |
Herrmann T, Große-Hovest L, Otz T, et al. (2008) Construction of optimized bispecific antibodies for selective activation of the death receptor CD95. Cancer Res 68: 1221-1227. doi: 10.1158/0008-5472.CAN-07-6175
|
| [28] |
Kaplan DH, Shankaran V, Dighe AS, et al. (1998) Demonstration of an interferon γ-dependent tumor surveillance system in immunocompetent mice. Proc Natl Acad Sci USA 95: 7556-7561. doi: 10.1073/pnas.95.13.7556
|
| [29] |
Gravett AM, Dalgleish AG, Copier J (2019) In vitro culture with gemcitabine augments death receptor and NKG2D ligand expression on tumour cells. Sci Rep 9: 1544. doi: 10.1038/s41598-018-38190-2
|
| [30] |
Micheau O, Solary E, Hammann A, et al. (1997) Sensitization of cancer cells treated with cytotoxic drugs to Fas-mediated cytotoxicity. J Natl Cancer Inst 89: 783-789. doi: 10.1093/jnci/89.11.783
|
| [31] |
Micheau O, Solary E, Hammann A, et al. (1999) Fas ligand-independent, FADD-mediated activation of the Fas death pathway by anticancer drugs. J Biol Chem 274: 7987-7992. doi: 10.1074/jbc.274.12.7987
|
| [32] |
Micheau O, Hammann A, Solary E, et al. (1999) STAT-1-independent upregulation of FADD and procaspase-3 and -8 in cancer cells treated with cytotoxic drugs. Biochem Biophys Res Comm 256: 603-607. doi: 10.1006/bbrc.1999.0391
|
| [33] | Petak I, Tillman DM, Houghton JA (2000) p53 dependence of Fas induction and acute apoptosis in response to 5-fluorouracil-leucovorin in human colon carcinoma cell lines. Clin Cancer Res 6: 4432-4441. |
| [34] |
Bergmann-Leitner ES, Abrams SI (2001) Treatment of human colon cell lines with anti-neoplastic agents enhances their lytic sensitivity to antigen-specific CD8+ cytotoxic T lymphocytes. Cancer Immunol Immunother 50: 445-455. doi: 10.1007/s002620100229
|
| [35] |
van Geelen CMM, de Vries EGE, Le TKP, et al. (2003) Differential modulation of the TRAIL receptors and the CD95 receptor in colon carcinoma cell lines. Br J Cancer 89: 363-373. doi: 10.1038/sj.bjc.6601065
|
| [36] |
Lacour S, Micheau O, Hammann A, et al. (2003) Chemotherapy enhances TNF-related apoptosis-inducing ligand DISC assembly in HT29 human colon cancer cells. Oncogene 22: 1807-1816. doi: 10.1038/sj.onc.1206127
|
| [37] |
Dong YB, Yang HL, McMasters KM (2003) E2F-1 overexpression sensitizes colorectal cancer cells to camptothecin. Cancer Gene Ther 10: 168-178. doi: 10.1038/sj.cgt.7700565
|
| [38] |
Lacour S, Hammann A, Grazide S, et al. (2004) Cisplatin-induced CD95 redistribution into membrane lipid rafts of HT29 human colon cancer cells. Cancer Res 64: 3593-3598. doi: 10.1158/0008-5472.CAN-03-2787
|
| [39] |
McDermott U, Longley DB, Galligan L, et al. (2005) Effect of p53 status and STAT1 on chemotherapy-induced, Fas-mediated apoptosis in colorectal cancer. Cancer Res 65: 8951-8960. doi: 10.1158/0008-5472.CAN-05-0961
|
| [40] |
Lin HH, Shi MD, Tseng HC, et al. (2014) Andrographolide sensitizes the cytotoxicity of human colorectal carcinoma cells toward cisplatin via enhancing apoptosis pathways in vitro and in vivo. Toxicol Sci 139: 108-120. doi: 10.1093/toxsci/kfu032
|
| [41] |
Pace E, Melis M, Siena L, et al. (2000) Effects of gemcitabine on cell proliferation and apoptosis in non-small-cell lung cancer (NSCLC) cell lines. Cancer Chemother Pharmacol 46: 467-476. doi: 10.1007/s002800000183
|
| [42] |
Supino R, Perego P, Gatti L, et al. (2001) A role for c-myc in DNA damage-induced apoptosis in a human TP53-mutant small-cell lung cancer cell line. Eur J Cancer 37: 2247-2256. doi: 10.1016/S0959-8049(01)00268-4
|
| [43] |
Okouoyo S, Herzer K, Ucur E, et al. (2004) Rescue of death receptor and mitochondrial apoptosis signaling in resistant human NSCLC in vivo. Int J Cancer 108: 580-587. doi: 10.1002/ijc.11585
|
| [44] |
Yoshimoto Y, Kawada M, Ikeda D, et al. (2005) Involvement of doxorubicin-induced Fas expression in the antitumor effect of doxorubicin on Lewis lung carcinoma in vivo. Int Immunopharmacol 5: 281-288. doi: 10.1016/j.intimp.2004.09.032
|
| [45] | Li Y, Xing D, Chen Q, et al. (2010) Enhancement of chemotherapeutic agent-induced apoptosis by inhibition of NF-κB using ursolic acid. Int J Cancer 127: 462-473. |
| [46] |
Siena L, Pace E, Ferraro M, et al. (2013) Gemcitabine sensitizes lung cancer cells to Fas/FasL system-mediated killing. Immunology 141: 242-255. doi: 10.1111/imm.12190
|
| [47] |
Wang LH, Li Y, Yang SN, et al. (2014) Gambogic acid synergistically potentiates cisplatin-induced apoptosis in non-small-cell lung cancer through suppressing NF-κB and MAPK/HO-1 signaling. Br J Cancer 110: 341-352. doi: 10.1038/bjc.2013.752
|
| [48] |
Weller M, Winter S, Schmidt C, et al. (1997) Topoisomerase-I inhibitors for human malignant glioma: differential modulation of p53, p21, bax, and bcl-2 expression and of CD95-mediated apoptosis by camptothecin and β-lapachone. Int J Cancer 73: 707-714. doi: 10.1002/(SICI)1097-0215(19971127)73:5<707::AID-IJC16>3.0.CO;2-2
|
| [49] |
Fulda S, Lutz W, Schwab M, et al. (1999) MycN sensitizes neuroblastoma cells for drug-induced apoptosis. Oncogene 18: 1479-1486. doi: 10.1038/sj.onc.1202435
|
| [50] |
Ciusani E, Perego P, Carenini N, et al. (2002) Fas/CD95-mediated apoptosis in human glioblastoma cells: a target for sensitization to topoisomerase I inhibitors. Biochem Pharmacol 63: 881-887. doi: 10.1016/S0006-2952(01)00837-1
|
| [51] |
Bian X, Giordano TD, Lin HJ, et al. (2004) Chemotherapy-induced apoptosis of S-type neuroblastoma cells requires caspase-9 and is augmented by CD95/Fas stimulation. J Biol Chem 279: 4663-4669. doi: 10.1074/jbc.M306905200
|
| [52] |
Terrasson J, Allart S, Martin H, et al. (2005) p73-dependent apoptosis through death receptor: impairment by human cytomegalovirus infection. Cancer Res 65: 2787-2794. doi: 10.1158/0008-5472.CAN-04-2019
|
| [53] |
Xia S, Rosen EM, Laterra J (2005) Sensitization of glioma cells to Fas-dependent apoptosis by chemotherapy-induced oxidative stress. Cancer Res 65: 5248-5255. doi: 10.1158/0008-5472.CAN-04-4332
|
| [54] |
Galenkamp KMO, Carriba P, Urresti J, et al. (2015) TNFα sensitizes neuroblastoma cells to FasL-, cisplatin- and etoposide-induced cell death by NF-κB-mediated expression of Fas. Mol Cancer 14: 62. doi: 10.1186/s12943-015-0329-x
|
| [55] |
Uslu R, Jewett A, Bonavida B (1996) Sensitization of human ovarian tumor cells by subtoxic CDDP to anti-Fas antibody-mediated cytotoxicity and apoptosis. Gynecol Oncol 62: 282-291. doi: 10.1006/gyno.1996.0228
|
| [56] | Uslu R, Borsellino N, Frost P, et al. (1997) Chemosensitization of human prostate carcinoma cell lines to anti-Fas-mediated cytotoxicity and apoptosis. Clin Cancer Res 3: 963-972. |
| [57] |
Mansouri A, Ridgway LD, Korapati A, et al. (2003) Sustained activation of JNK/p38 MAPK pathways in response to cisplatin leads to Fas ligand induction and cell death in ovarian carcinoma cells. J Biol Chem 278: 19245-19256. doi: 10.1074/jbc.M208134200
|
| [58] |
Bagnoli M, Balladore E, Luison E, et al. (2007) Sensitization of p53-mutated epithelial ovarian cancer to CD95-mediated apoptosis is synergistically induced by cisplatin pretreatment. Mol Cancer Ther 6: 762-772. doi: 10.1158/1535-7163.MCT-06-0357
|
| [59] |
Alagkiozidis I, Facciabene A, Carpenito C, et al. (2009) Increased immunogenicity of surviving tumor cells enables cooperation between liposomal doxorubicin and IL-18. J Transl Med 7: 104. doi: 10.1186/1479-5876-7-104
|
| [60] |
Karaca B, Atmaca H, Bozkurt E, et al. (2013) Combination of AT-101/cisplatin overcomes chemoresistance by inducing apoptosis and modulating epigenetics in human ovarian cancer cells. Mol Biol Rep 40: 3925-3933. doi: 10.1007/s11033-012-2469-z
|
| [61] |
Frost P, Ng CP, Belldegrun A, et al. (1997) Immunosensitization of prostate carcinoma cell lines for lymphocytes (CTL, TIL, LAK)-mediated apoptosis via the Fas-Fas-ligand pathway of cytotoxicity. Cell Immunol 180: 70-83. doi: 10.1006/cimm.1997.1169
|
| [62] |
Costa-Pereira AP, Cotter TG (1999) Camptothecin sensitizes androgen-independent prostate cancer cells to anti-Fas-induced apoptosis. Br J Cancer 80: 371-378. doi: 10.1038/sj.bjc.6690365
|
| [63] |
Costa-Pereira AP, McKenna SL, Cotter TG (2000) Activation of SAPK/JNK by camptothecin sensitizes androgen-independent prostate cancer cells to Fas-induced apoptosis. Br J Cancer 82: 1827-1834. doi: 10.1054/bjoc.2000.1149
|
| [64] |
Frost PJ, Butterfield LH, Dissette VB, et al. (2001) Immunosensitization of melanoma tumor cells to non-MHC Fas-mediated killing by MART-1-specific CTL cultures. J Immunol 166: 3564-3573. doi: 10.4049/jimmunol.166.5.3564
|
| [65] |
Das A, Durrant D, Mitchell C, et al. (2015) Sildenafil (Viagra) sensitizes prostate cancer cells to doxorubicin-mediated apoptosis through CD95. Oncotarget 7: 4399-4413. doi: 10.18632/oncotarget.6749
|
| [66] |
Ruiz-Ruiz MC, López-Rivas A (1999) p53-mediated up-regulation of CD95 is not involved in genotoxic drug-induced apoptosis of human breast tumor cells. Cell Death Differ 6: 271-280. doi: 10.1038/sj.cdd.4400490
|
| [67] | Ruiz-Ruiz C, Muñoz-Pinedo C, López-Rivas A (2000) Interferon-γ treatment elevates caspase-8 expression and sensitizes human breast tumor cells to a death receptor-induced mitochondria-operated apoptotic program. Cancer Res 60: 5673-5680. |
| [68] |
Basma H, El-Refaey H, Sgagias MK, et al. (2005) Bcl-2 antisense and cisplatin combination treatment of MCF-7 breast cancer cells with or without functional p53. J Biomed Sci 12: 999-1011. doi: 10.1007/s11373-005-9025-y
|
| [69] |
Mohammad N, Singh SV, Malvi P, et al. (2015) Strategy to enhance efficacy of doxorubicin in solid tumor cells by methyl-β-cyclodextrin: involvement of p53 and Fas receptor ligand complex. Sci Rep 5: 11853. doi: 10.1038/srep11853
|
| [70] |
Posovszky C, Friesen C, Herr I, et al. (1999) Chemotherapeutic drugs sensitize pre-B ALL cells for CD95- and cytotoxic T-lymphocyte-mediated apoptosis. Leukemia 13: 400-409. doi: 10.1038/sj.leu.2401327
|
| [71] |
Ortiz-Lazareno PC, Bravo-Cuellar A, Lerma-Díaz JM, et al. (2014) Sensitization of U937 leukemia cells to doxorubicin by the MG132 proteasome inhibitor induces an increase in apoptosis by suppressing NF-kappa B and mitochondrial membrane potential loss. Cancer Cell Int 14: 13. doi: 10.1186/1475-2867-14-13
|
| [72] |
Duverger V, Sartorius U, Klein-Bauernschmitt P, et al. (2002) Enhancement of cisplatin-induced apoptosis by infection with adeno-associated virus type 2. Int J Cancer 97: 706-712. doi: 10.1002/ijc.10077
|
| [73] |
Lim YS, So HS, Kim MS, et al. (2002) Palgin sensitizes the Adriamycin-induced apoptosis via the enhancement of Fas/Fas ligand expression. Life Sci 71: 2391-2401. doi: 10.1016/S0024-3205(02)02039-8
|
| [74] |
Hougardy BMT, van der Zee AGJ, van den Heuvel FAJ, et al. (2005) Sensitivity to Fas-mediated apoptosis in high-risk HPV-positive human cervical cancer cells: relationship with Fas, caspase-8, and Bid. Gynecol Oncol 97: 353-364. doi: 10.1016/j.ygyno.2005.01.036
|
| [75] | Sui Y, Yang Y, Wang J, et al. (2015) Lysophosphatidic acid inhibits apoptosis induced by cisplatin in cervical cancer cells. BioMed Res Int 2015: 598386. |
| [76] |
Kinoshita H, Yoshikawa H, Shiiki K, et al. (2000) Cisplatin (CDDP) sensitizes human osteosarcoma cell to Fas/CD95-mediated apoptosis by down-regulating FLIP-L expression. Int J Cancer 88: 986-991. doi: 10.1002/1097-0215(20001215)88:6<986::AID-IJC23>3.0.CO;2-B
|
| [77] |
Yuan XW, Zhu XF, Huang XF, et al. (2007) p14ARF sensitizes human osteosarcoma cells to cisplatin-induced apoptosis in a p53-independent manner. Cancer Biol Ther 6: 1074-1080. doi: 10.4161/cbt.6.7.4324
|
| [78] |
Huang T, Gong WH, Zou CP, et al. (2014) Marsdenia tenacissima extract sensitizes MG63 cells to doxorubicin-induced apoptosis. Genet Mol Res 13: 354-362. doi: 10.4238/2014.January.21.3
|
| [79] |
Pei Q, Pan J, Ding X, et al. (2015) Gemcitabine sensitizes pancreatic cancer cells to the CTLs antitumor response induced by BCG-stimulated dendritic cells via a Fas-dependent pathway. Pancreatology 15: 233-239. doi: 10.1016/j.pan.2015.04.001
|
| [80] |
Pietkiewicz S, Eils R, Krammer PH, et al. (2015) Combinatorial treatment of CD95L and gemcitabine in pancreatic cancer cells induces apoptotic and RIP1-mediated necroptotic cell death network. Exp Cell Res 339: 1-9. doi: 10.1016/j.yexcr.2015.10.005
|
| [81] |
Yang S, Haluska FG (2004) Treatment of melanoma with 5-fluorouracil or dacarbazine in vitro sensitizes cells to antigen-specific CTL lysis through perforin/granzyme- and Fas-mediated pathways. J Immunol 172: 4599-4608. doi: 10.4049/jimmunol.172.7.4599
|
| [82] | Wu XX, Mizutani Y, Kakehi Y, et al. (2000) Enhancement of Fas-mediated apoptosis in renal cell carcinoma cells by adriamycin. Cancer Res 60: 2912-2818. |
| [83] |
Kuwahara D, Tsutsumi K, Kobayashi T, et al. (2000) Caspase-9 regulates cisplatin-induced apoptosis in human head and neck squamous cell carcinoma cells. Cancer Lett 148: 65-71. doi: 10.1016/S0304-3835(99)00315-8
|
| [84] |
Kataoka T, Ito M, Budd RC, et al. (2002) Expression level of c-FILP versus Fas determines susceptibility to Fas ligand-induced cell death in murine thymoma EL-4 cells. Exp Cell Res 273: 256-264. doi: 10.1006/excr.2001.5438
|
| [85] |
Iwase M, Watanabe H, Kondo G, et al. (2003) Enhanced susceptibility of oral squamous cell carcinoma cell lines to Fas-mediated apoptosis by cisplatin and 5-fluorouracil. Int J Cancer 106: 619-625. doi: 10.1002/ijc.11239
|
| [86] |
Spierings DCJ, de Vries EGE, Stel AJ, et al. (2004) Low p21Waf1/Cip1 protein level sensitizes testicular germ cell tumor cells to Fas-mediated apoptosis. Oncogene 23: 4862-4872. doi: 10.1038/sj.onc.1207617
|
| [87] |
Yamana K, Bilim V, Hara N, et al. (2005) Prognostic impact of Fas/CD95/APO-1 in urothelial cancers: decreased expression of Fas is associated with disease progression. Br J Cancer 93: 544-551. doi: 10.1038/sj.bjc.6602732
|
| [88] |
Nitobe J, Yamaguchi S, Okuyama M, et al. (2003) Reactive oxygen species regulate FLICE inhibitory protein (FLIP) and susceptibility to Fas-mediated apoptosis in cardiac myocytes. Cardiovasc Res 57: 119-128. doi: 10.1016/S0008-6363(02)00646-6
|
| [89] |
Labroille G, Dumain P, Lacombe F, et al. (2000) Flow cytometric evaluation of fas expression in relation to response and resistance to anthracyclines in leukemic cells. Cytometry 39: 195-202. doi: 10.1002/(SICI)1097-0320(20000301)39:3<195::AID-CYTO4>3.0.CO;2-A
|
| [90] |
Fulda S, Küfer MU, Meyer E, et al. (2001) Sensitization for death receptor- or drug-induced apoptosis by re-expression of caspase-8 through demethylation or gene transfer. Oncogene 20: 5865-5877. doi: 10.1038/sj.onc.1204750
|
| [91] |
Yang D, Torres CM, Bardhan K, et al. (2012) Decitabine and vorinostat cooperate to sensitize colon carcinoma cells to Fas ligand-induced apoptosis in vitro and tumor suppression in vivo. J Immunol 188: 4441-4449. doi: 10.4049/jimmunol.1103035
|
| [92] |
Mishima K, Nariai Y, Yoshimura Y (2003) Carboplatin induces Fas (APO-1/CD95)-dependent apoptosis of human tongue carcinoma cells: sensitization for apoptosis by upregulation of FADD expression. Int J Cancer 105: 593-600. doi: 10.1002/ijc.11133
|
| [93] |
Woo SH, Park IC, Park MJ, et al. (2004) Arsenic trioxide sensitizes CD95/Fas-induced apoptosis through ROS-mediated upregulation of CD95/Fas by NF-kappaB activation. Int J Cancer 112: 596-606. doi: 10.1002/ijc.20433
|
| [94] |
Hallett WH, Ames E, Motarjemi M, et al. (2008) Sensitization of tumor cells to NK cell-mediated killing by proteasome inhibition. J Immunol 180: 163-170. doi: 10.4049/jimmunol.180.1.163
|
| [95] |
Symes JC, Kunin M, Fleshner NE, et al. (2008) Fas-mediated killing of primary prostate cancer cells is increased by mitoxantrone and docetaxel. Mol Cancer Ther 7: 3018-3028. doi: 10.1158/1535-7163.MCT-08-0335
|
| [96] |
Llobet D, Eritja N, Yeramian A, et al. (2010) The multikinase inhibitor sorafenib induces apoptosis and sensitizes endometrial cancer cells to TRAIL by different mechanisms. Eur J Cancer 46: 836-850. doi: 10.1016/j.ejca.2009.12.025
|
| [97] |
Zhang G, Park MA, Mitchell C, et al. (2008) Vorinostat and sorafenib synergistically kill tumor cells via FLIP suppression and CD95 activation. Clin Cancer Res 14: 5385-5399. doi: 10.1158/1078-0432.CCR-08-0469
|
| [98] |
Bonnotte B, Favre N, Reveneau S, et al. (1998) Cancer cell sensitization to Fas-mediated apoptosis by sodium butyrate. Cell Death Differ 5: 480-487. doi: 10.1038/sj.cdd.4400371
|
| [99] |
Rivkin I, Cohen K, Bod T, et al. (2014) Cancer cell sensitization and improved treatment efficacy by combined sodium butyrate and paclitaxel formulations is cancer-type specific. Int J Pharm 461: 437-447. doi: 10.1016/j.ijpharm.2013.12.021
|
| [100] |
Kondo G, Iwase M, Watanabe H, et al. (2006) Enhancement of susceptibility to Fas-mediated apoptosis in oral squamous cell carcinoma cells by phosphatidylinositol 3-kinase inhibitor. Oral Oncol 42: 745-752. doi: 10.1016/j.oraloncology.2005.11.015
|
| [101] |
Rao-Bindal K, Zhou Z, Kleinerman ES (2012) MS-275 sensitizes osteosarcoma cells to Fas ligand-induced cell death by increasing the localization of Fas in membrane lipid rafts. Cell Death Dis 3: e369. doi: 10.1038/cddis.2012.101
|
| [102] |
Castro BM, de Almeida RF, Goormaghtigh E, et al. (2011) Organization and dynamics of Fas transmembrane domain in raft membranes and modulation by ceramide. Biophys J 101: 1632-1641. doi: 10.1016/j.bpj.2011.08.022
|
| [103] |
Gajate C, Del Canto-Jañez E, Acuña AU, et al. (2004) Intracellular triggering of Fas aggregation and recruitment of apoptotic molecules into Fas-enriched rafts in selective tumor cell apoptosis. J Exp Med 200: 353-365. doi: 10.1084/jem.20040213
|
| [104] |
Mollinedo F, de la Iglesia-Vicente J, Gajate C, et al. (2010) In vitro and in vivo selective antitumor activity of Edelfosine against mantle cell lymphoma and chronic lymphocytic leukemia involving lipid rafts. Clin Cancer Res 16: 2046-2054. doi: 10.1158/1078-0432.CCR-09-2456
|
| [105] |
Rajesh D, Stenzel RA, Howard SP (2003) Perillyl alcohol as a radio-/chemosensitizer in malignant glioma. J Biol Chem 278: 35968-35978. doi: 10.1074/jbc.M303280200
|
| [106] |
Westerndorp MO, Frank R, Ochsenbauer C, et al. (1995) Sensitization of T cells to CD95-mediated apoptosis by HIV-1 Tat and gp120. Nature 375: 497-500. doi: 10.1038/375497a0
|
| [107] |
Ruggieri A, Harada T, Matsuura Y, et al. (1997) Sensitization to Fas-mediated apoptosis by hepatitis C virus core protein. Virology 229: 68-76. doi: 10.1006/viro.1996.8420
|
| [108] | Tanaka M, Suda T, Yatomi T, et al. (1997) Lethal effect of recombinant human Fas ligand in mice pretreated with Propionibacterium acnes. J Immunol 158: 2303-2309. |
| [109] |
Wagner S, Beil W, Westermann J, et al. (1997) Regulation of gastric epithelial cell growth by Helicobacter pylori: evidence for a major role of apoptosis. Gastroenterology 113: 1836-1847. doi: 10.1016/S0016-5085(97)70003-9
|
| [110] |
Chakraborty M, Abrams SI, Camphausen K, et al. (2003) Irradiation of tumor cells up-regulates Fas and enhances CTL lytic activity and CTL adoptive immunotherapy. J Immunol 170: 6338-6347. doi: 10.4049/jimmunol.170.12.6338
|
| [111] | Park IC, Woo SH, Park MJ, et al. (2003) Ionizing radiation and nitric oxide donor sensitize Fas-induced apoptosis via up-regulation of Fas in human cervical cancer cells. Oncol Rep 10: 629-633. |
| [112] |
Schroder K, Herztog PJ, Ravasi T, et al. (2004) Interferon-γ: an overview of signals, mechanisms and functions. J Leukoc Biol 75: 163-189. doi: 10.1189/jlb.0603252
|
| [113] |
Zaidi MR, Merlino G (2011) The two faces of interferon-γ in cancer. Clin Cancer Res 17: 6118-6124. doi: 10.1158/1078-0432.CCR-11-0482
|
| [114] |
Meissl K, Macho-Maschler S, Müller M, et al. (2015) The good and the bad faces of STAT1 in solid tumors. Cytokine 89: 12-20. doi: 10.1016/j.cyto.2015.11.011
|
| [115] |
Showalter A, Limaye A, Oyer JL, et al. (2017) Cytokines in immunogenic cell death: applications for cancer immunotherapy. Cytokine 97: 123-132. doi: 10.1016/j.cyto.2017.05.024
|
| [116] |
Burke JD, Young HA (2019) IFN-γ: a cytokine at the right time, is in the right place. Semin Immunol 43: 101280. doi: 10.1016/j.smim.2019.05.002
|
| [117] |
Yonehara S, Ishii A, Yonehara M (1989) A cell-killing monoclonal antibody (anti-Fas) to a cell surface antigen co-downregulated with the receptor of tumor necrosis factor. J Exp Med 169: 1747-1756. doi: 10.1084/jem.169.5.1747
|
| [118] |
Günthert AR, Sträter J, von Reyher U, et al. (1996) Early detachment of colon carcinoma cells during CD95 (APO-1/Fas)-mediated apoptosis. I. de-adhesion from hyaluronate by shedding of CD44. J Cell Biol 134: 1089-1096. doi: 10.1083/jcb.134.4.1089
|
| [119] |
Ossina NK, Cannas A, Powers VC, et al. (1997) Interferon-γ modulates a p53-independent apoptotic pathway and apoptosis-related gene expression. J Biol Chem 272: 16351-16357. doi: 10.1074/jbc.272.26.16351
|
| [120] |
Tillman DM, Harwood FG, Gibson AA, et al. (1998) Expression of genes that regulate Fas signaling and Fas-mediated apoptosis in colon carcinoma cells. Cell Death Differ 5: 450-457. doi: 10.1038/sj.cdd.4400369
|
| [121] |
Darcy PK, Kershaw MH, Trapani JA, et al. (1998) Expression in cytotoxic T lymphocytes of a single-chain anti-carcinoembryonic antigen antibody. Redirected Fas ligand-mediated lysis of colon carcinoma. Eur J Immunol 28: 1663-1672. doi: 10.1002/(SICI)1521-4141(199805)28:05<1663::AID-IMMU1663>3.0.CO;2-L
|
| [122] |
Koshiji M, Adachi Y, Sogo S, et al. (1998) Apoptosis of colorectal adenocarcinoma (COLO 201) by tumour necrosis factor (TNF-α) and/or interferon-gamma (IFN-γ), resulting from down-modulation of Bcl-2 expression. Clin Exp Immunol 111: 211-218. doi: 10.1046/j.1365-2249.1998.00460.x
|
| [123] | von Reyher U, Sträter J, Kittstein W, et al. (1998) Colon carcinoma cells use different mechanisms to escape CD95-mediated apoptosis. Cancer Res 58: 526-534. |
| [124] | Xu X, Fu XY, Plate J, et al. (1998) IFN-γ induces cell growth inhibition by Fas-mediated apoptosis: requirement of STAT1 protein for up-regulation of Fas and FasL expression. Cancer Res 58: 2832-2837. |
| [125] |
Rapoport E, Pendu JL (1999) Glycosylation alterations of cells in late phase apoptosis from colon carcinomas. Glycobiology 9: 1337-1345. doi: 10.1093/glycob/9.12.1337
|
| [126] | Tillman DM, Petak I, Houghton JA (1999) A Fas-dependent component in 5-fluorouracil/leucovorin-induced cytotoxicity in colon carcinoma cells. Clin Cancer Res 5: 425-430. |
| [127] |
Bergmann-Leitner ES, Abrams SI (2000) Influence of interferon γ on modulation of Fas expression by human colon carcinoma cells and their subsequent sensitivity to antigen-specific CD8+ cytotoxic T lymphocyte attack. Cancer Immunol Immunother 49: 193-207. doi: 10.1007/s002620000105
|
| [128] |
Bergmann-Leitner ES, Abrams SI (2000) Differential role of Fas/Fas ligand interactions in cytolysis of primary and metastatic colon carcinoma cell lines by human antigen-specific CD8+ CTL. J Immunol 164: 4941-4954. doi: 10.4049/jimmunol.164.9.4941
|
| [129] |
O'connell J, Bennett MW, Nally K, et al. (2000) Interferon-γ sensitizes colonic epithelial cell lines to physiological and therapeutic inducers of colonocyte apoptosis. J Cell Physiol 185: 331-338. doi: 10.1002/1097-4652(200012)185:3<331::AID-JCP3>3.0.CO;2-V
|
| [130] | Remacle-Bonnet MM, Garrouste FL, Heller S, et al. (2000) Insulin-like growth factor-I protects colon cancer cells from death factor-induced apoptosis by potentiating tumor necrosis factor α-induced mitogen-activated protein kinase and nuclear factor κB signaling pathways. Cancer Res 60: 2007-2017. |
| [131] | Manos EJ, Jones DA (2001) Assessment of tumor necrosis factor receptor and Fas signaling pathways by transcriptional profiling. Cancer Res 61: 433-438. |
| [132] |
Martin CA, Panja A (2002) Cytokine regulation of human intestinal primary epithelial cell susceptibility to Fas-mediated apoptosis. Am J Physiol Gastrointest Liver Physiol 282: G92-G104. doi: 10.1152/ajpgi.2002.282.1.G92
|
| [133] | Schwartzberg LS, Petak I, Stewart C, et al. (2002) Modulation of Fas signaling pathway by IFN-γ in therapy of colon cancer: phase I trial and correlative studies of IFN-γ, 5-fluorouracil, and leucovorin. Clin Cancer Res 8: 2488-2498. |
| [134] |
Wilson CA, Browning JL (2002) Death of HT29 adenocarcinoma cells induced by TNF family receptor activation is caspase-independent and displays features of both apoptosis and necrosis. Cell Death Differ 9: 1321-1333. doi: 10.1038/sj.cdd.4401107
|
| [135] | Geller J, Petak I, Szucs KS, et al. (2003) Interferon-γ-induced sensitization of colon carcinomas to ZD9331 targets caspases, downstream of Fas, independent of mitochondrial signaling and the inhibitor of apoptosis survivin. Clin Cancer Res 9: 6504-6515. |
| [136] |
Liu K, Abrams SI (2003) Coordinate regulation of IFN consensus sequence-binding protein and caspase-1 in the sensitization of human colon carcinoma cells to Fas-mediated apoptosis by IFN-γ. J Immunol 170: 6329-6337. doi: 10.4049/jimmunol.170.12.6329
|
| [137] |
Liu K, McDuffie E, Abrams SI (2003) Exposure of human primary colon carcinoma cells to anti-Fas interactions influences the emergence of pre-existing Fas-resistant metastatic subpopulations. J Immunol 171: 4164-4174. doi: 10.4049/jimmunol.171.8.4164
|
| [138] |
Seidelin JB, Jäättelä M, Nielsen OH (2004) Continuous interferon-γ or tumor necrosis factor-α exposure of enterocytes attenuates cell death responses. Cytokine 27: 113-119. doi: 10.1016/j.cyto.2004.04.001
|
| [139] |
Turner PK, Houghton JA, Petak I, et al. (2004) Interferon-gamma pharmacokinetics and pharmacodynamics in patients with colorectal cancer. Cancer Chemother Pharmacol 53: 253-260. doi: 10.1007/s00280-003-0723-8
|
| [140] |
Vekemans K, Braet F, Muyllaert D, et al. (2004) Nitric oxide from rat liver sinusoidal endothelial cells induces apoptosis in IFN γ-sensitized CC531s colon carcinoma cells. J Hepatol 41: 11-18. doi: 10.1016/j.jhep.2004.03.026
|
| [141] |
Siegmund D, Wicovsky A, Schmitz I (2005) Death receptor-induced signaling pathways are differentially regulated by gamma interferon upstream of caspase-8 processing. Mol Cell Biol 25: 6363-6379. doi: 10.1128/MCB.25.15.6363-6379.2005
|
| [142] | Saha A, Chatterjee SK, Foon KA, et al. (2006) Anti-idiotype antibody induced cellular immunity in mice transgenic for human carcinoembryonic antigen. Immunology 118: 483-496. |
| [143] |
Geng L, Zhu B, Dai BH, et al. (2011) A let-7/Fas double-negative feedback loop regulates human colon carcinoma cells sensitivity to Fas-related apoptosis. Biochem Biophys Res Comm 408: 494-499. doi: 10.1016/j.bbrc.2011.04.074
|
| [144] |
Shadrin N, Shapira MG, Khalfin B, et al. (2015) Serine protease inhibitors interact with IFN-γ through up-regulation of FasR; a novel therapeutic strategy against cancer. Exp Cell Res 330: 233-239. doi: 10.1016/j.yexcr.2014.11.005
|
| [145] |
Muraki M, Hirota K (2018) Confirmation of covalently-linked structure and cell-death inducing activity in site-specific chemical conjugates of human Fas ligand extracellular domain. BMC Res Notes 11: 395. doi: 10.1186/s13104-018-3501-8
|
| [146] | Muraki M, Hirota K (2019) Site-specific biotin-group conjugate of human Fas ligand extracellular domain: preparation and characterization of cell-death-inducing activity. Curr Top Pep Prot Res 20: 17-24. |
| [147] | Morimoto H, Yonehara S, Bonavida B (1993) Overcoming tumor necrosis factor and drug resistance of human tumor cell lines by combination treatment with anti-Fas antibody and drugs or toxins. Cancer Res 53: 2591-2596. |
| [148] |
Biswas P, Poli G, Orenstein JM, et al. (1994) Cytokine-mediated induction of human immunodeficiency virus (HIV) expression and cell death in chronically infected U1 cells: do tumor necrosis factor alpha and gamma interferon selectively kill HIV-infected cells? J Virol 68: 2598-2604. doi: 10.1128/JVI.68.4.2598-2604.1994
|
| [149] |
Shima Y, Nishimoto N, Ogata A, et al. (1995) Myeloma cells express Fas antigen/APO-1 (CD95) but only some are sensitive to anti-Fas antibody resulting in apoptosis. Blood 85: 757-764. doi: 10.1182/blood.V85.3.757.bloodjournal853757
|
| [150] |
Efferth T, Fabry U, Osieka R (1996) Anti-Fas/Apo-1 monoclonal antibody CH-11 depletes glutathione and kills multidrug-resistant human leukemic cells. Blood Cell Mol Dis 22: 2-9. doi: 10.1006/bcmd.1996.0002
|
| [151] |
Spets H, Georgii-Hemming P, Siljason J, et al. (1998) Fas/APO-1 (CD95)-mediated apoptosis is activated by interferon-γ and interferon-α in interleukin-6 (IL-6)-dependent and IL-6-independent multiple myeloma cell lines. Blood 92: 2914-2923. doi: 10.1182/blood.V92.8.2914
|
| [152] |
Horie T, Dobashi K, Iizuka K, et al. (1999) Interferon-γ rescues TNF-α-induced apoptosis mediated by up-regulation of TNFR2 on EoL-1 cells. Exp Hematol 27: 512-519. doi: 10.1016/S0301-472X(98)00058-7
|
| [153] |
Sata M, Suhara T, Walsh K (2000) Vascular endothelial cells and smooth muscle cells differ in expression of Fas and Fas ligand and in sensitivity to Fas ligand-induced cell death. Implications for vascular disease and therapy. Arterioscler Thromb Vasc Biol 20: 309-316. doi: 10.1161/01.ATV.20.2.309
|
| [154] |
Varela N, Muñoz-Pinedo C, Ruiz-Ruiz C, et al. (2001) Interferon-γ sensitizes human myeloid leukemia cells to death receptor-mediated apoptosis by a pleiotropic mechanism. J Biol Chem 276: 17779-17787. doi: 10.1074/jbc.M100815200
|
| [155] |
Dörrie J, Sapala K, Zunino SJ (2002) Interferon-γ increases the expression of glycosylated CD95 in B-leukemic cells: an inducible model to study the role of glycosylation in CD95-signalling and trafficking. Cytokine 18: 98-107. doi: 10.1006/cyto.2002.1030
|
| [156] |
Jedema I, Barge RMY, Willemze R, et al. (2003) High susceptibility of human leukemic cells to Fas-induced apoptosis is restricted to G1 phase of the cell cycle and can be increased by interferon treatment. Leukemia 17: 576-584. doi: 10.1038/sj.leu.2402844
|
| [157] |
Dimberg LY, Dimberg AI, Ivarsson K, et al. (2005) Ectopic and IFN-induced expression of Fas overcomes resistance to Fas-mediated apoptosis in multiple myeloma cells. Blood 106: 1346-1354. doi: 10.1182/blood-2004-04-1322
|
| [158] |
Guy CS, Wang J, Michalak TI (2006) Hepatocytes as cytotoxic effector cells can induce cell death by CD95 ligand-mediated pathway. Hepatology 43: 1231-1240. doi: 10.1002/hep.21201
|
| [159] |
Boselli D, Ragimbeau J, Orlando L, et al. (2010) Expression of IFNγR2 mutated in a dileucine internalization motif reinstates IFNγ signaling and apoptosis in human T lymphocytes. Immunol Lett 134: 17-25. doi: 10.1016/j.imlet.2010.08.005
|
| [160] |
Dimberg LY, Dimberg A, Ivarsson K, et al. (2012) Stat1 activation attenuates IL-6 induced Stat3 activity but does not alter apoptosis sensitivity in multiple myeloma. BMC Cancer 12: 318. doi: 10.1186/1471-2407-12-318
|
| [161] |
Fujihara Y, Takato T, Hoshi K (2014) Macrophage-inducing FasL on chondrocytes forms immune privilege in cartilage tissue engineering, enhancing in vivo regeneration. Stem Cell 32: 1208-1219. doi: 10.1002/stem.1636
|
| [162] | Xia HL, Li CJ, Hou XF, et al. (2017) Interferon-γ affects leukemia cell apoptosis through regulating Fas/FasL signaling pathway. Eur Rev Med Pharmacol Sci 21: 2244-2248. |
| [163] |
Weller M, Frei K, Groscurth P, et al. (1994) Anti-Fas/APO-1 antibody-mediated apoptosis of cultured human glioma cells. Induction and modulation of sensitivity by cytokines. J Clin Invest 94: 954-964. doi: 10.1172/JCI117462
|
| [164] |
Fulda S, Debatin KM (2002) IFNγ sensitizes for apoptosis by upregulating caspase-8 expression through the Stat1 pathway. Oncogene 21: 2295-2308. doi: 10.1038/sj.onc.1205255
|
| [165] |
Buntinx M, Gielen E, van Hummelen P, et al. (2004) Cytokine-induced cell death in human oligodendroglial cell lines. II: alterations in gene expression induced by interferon-γ and tumor necrosis factor-α. J Neurosci Res 76: 846-861. doi: 10.1002/jnr.20117
|
| [166] |
Choi C, Jeong E, Benveniste EN (2004) Caspase-1 mediates Fas-induced apoptosis and is up-regulated by interferon-γ in human astrocytoma cells. J Neuro-oncol 67: 167-176. doi: 10.1023/B:NEON.0000021896.52664.9e
|
| [167] |
Song JH, Wang CX, Song DK, et al. (2005) Interferon γ induces neurite outgrowth by up-regulation of p35 neuron-specific cycline-dependent kinase 5 activator via activation of ERK1/2 pathway. J Biol Chem 280: 12896-12901. doi: 10.1074/jbc.M412139200
|
| [168] |
Giammarioli AM, Vona R, Gambardella L, et al. (2009) Interferon-γ bolsters CD95/Fas-mediated apoptosis of astroglioma cells. FEBS J 276: 5920-5935. doi: 10.1111/j.1742-4658.2009.07271.x
|
| [169] | Wen LP, Madani K, Fahrni JA, et al. (1997) Dexamethasone inhibits lung epithelial cell apoptosis induced by IFN-γ and Fas. Am J Physiol 273: L921-L929. |
| [170] |
Maeyama T, Kuwano K, Kawasaki M, et al. (2001) Upregulation of Fas-signaling molecules in lung epithelial cells from patients with idiopathic pulmonary fibrosis. Eur Respir J 17: 180-189. doi: 10.1183/09031936.01.17201800
|
| [171] |
Coulter KR, Doseff A, Sweeney P, et al. (2002) Opposing effect by cytokines on Fas-mediated apoptosis in A549 lung epithelial cells. Am J Respir Cell Mol Biol 26: 58-66. doi: 10.1165/ajrcmb.26.1.4285
|
| [172] |
Kim KB, Choi YH, Kim IK, et al. (2002) Potentiation of Fas- and TRAIL-mediated apoptosis by IFN-γ in A549 lung epithelial cells: enhancement of caspase-8 expression through IFN-response element. Cytokine 20: 283-288. doi: 10.1006/cyto.2003.2008
|
| [173] |
Hopkins-Donaldson S, Ziegler A, Kurtz S, et al. (2003) Silencing of death receptor and caspase-8 expression in small cell lung carcinoma cell lines and tumours by DNA methylation. Cell Death Differ 10: 356-364. doi: 10.1038/sj.cdd.4401157
|
| [174] |
Crescenzi E, Pacifico F, Lavorgna A, et al. (2011) NF-κB-dependent cytokine secretion controls Fas expression on chemotherapy-induced premature senescent tumor cells. Oncogene 30: 2707-2717. doi: 10.1038/onc.2011.1
|
| [175] | Keane MM, Ettenberg SA, Lowrey GA, et al. (1996) Fas expression and function in normal and malignant breast cell lines. Cancer Res 56: 4791-4798. |
| [176] |
Haynes NM, Smyth MJ, Kershaw MH, et al. (1999) Fas-ligand-mediated lysis of erbB-2-expressing tumour cells by redirected cytotoxic T lymphocytes. Cancer Immunol Immunother 47: 278-286. doi: 10.1007/s002620050532
|
| [177] |
Danforth DN, Zhu Y (2005) Conversion of Fas-resistant to Fas-sensitive MCF-7 breast cancer cells by the synergistic interaction of interferon-γ and all-trans retinoic acid. Breast Cancer Res Treat 94: 81-91. doi: 10.1007/s10549-005-7491-6
|
| [178] |
Deiss LP, Galinka H, Beriss H, et al. (1996) Cathepsin D protease mediates programmed cell death induced by interferon-γ, Fas/APO-1 and TNF-α. EMBO J 15: 3861-3870. doi: 10.1002/j.1460-2075.1996.tb00760.x
|
| [179] |
Jauharoh SNA, Saegusa J, Sugimoto T, et al. (2012) SS-A/Ro52 promotes apoptosis by regulating Bcl-2 production. Biochem Biophys Res Comm 417: 582-587. doi: 10.1016/j.bbrc.2011.12.010
|
| [180] |
Boeddeker SJ, Baston-Buest DM, Altergot-Ahmad O, et al. (2014) Syndecan-1 knockdown in endometrial epithelial cells alters their apoptotic protein profile and enhances the inducibility of apoptosis. Mol Hum Reprod 20: 567-578. doi: 10.1093/molehr/gau009
|
| [181] |
Yang D, Thangaraju M, Browing DD, et al. (2007) IFN regulatory factor 8 mediates apoptosis in nonhemopoietic tumor cells via regulation of Fas expression. J Immunol 179: 4775-4782. doi: 10.4049/jimmunol.179.7.4775
|
| [182] |
Banik D, Khan ANH, Walseng E, et al. (2012) Interferon regulatory factor-8 is important for histone deacetylase inhibitor-mediated antitumor activity. Plos One 7: e45422. doi: 10.1371/journal.pone.0045422
|
| [183] |
Nonomura N, Miki T, Yokoyama M, et al. (1996) Fas/APO-1-mediated apoptosis of human renal cell carcinoma. Biochem Biophys Res Comm 229: 945-951. doi: 10.1006/bbrc.1996.1906
|
| [184] |
Lee JK, Sayers TJ, Brooks AD, et al. (2000) IFN-γ-dependent delay of in vivo tumor progression by Fas overexpression of murine renal cancer cells. J Immunol 164: 231-239. doi: 10.4049/jimmunol.164.1.231
|
| [185] |
Tomita Y, Bilim V, Hara N, et al. (2003) Role of IRF-1 and caspase-7 in IFN-γ enhancement of Fas-mediated apoptosis in ACHN renal cell carcinoma cells. Int J Cancer 104: 400-408. doi: 10.1002/ijc.10956
|
| [186] |
García-Sánchez O, López-Novoa JM, López-Hermández FJ (2014) Interferon-γ reduces the proliferation of primed human renal tubular cells. Nephron Extra 4: 1-7. doi: 10.1159/000353587
|
| [187] |
Ugurel S, Seiter S, Pappl G, et al. (1999) Heterogeneous susceptibility to CD95-induced apoptosis in melanoma cells correlates with bcl-2 and bcl-x expression and is sensitive to modulation by interferon-γ. Int J Cancer 82: 727-736. doi: 10.1002/(SICI)1097-0215(19990827)82:5<727::AID-IJC17>3.0.CO;2-E
|
| [188] |
Shchors K, Yehiely F, Deiss LP (2004) Cell death inhibiting RNA (CDIR) modulates IFN-γ-stimulated sensitization to Fas/CD95/Apo-1 and TRAIL/Apo2L-induced apoptosis. Cell Cycle 3: 1606-1611. doi: 10.4161/cc.3.12.1295
|
| [189] | Hiramoto K, Inui M, Kamei T, et al. (2006) mHFE7A, a newly identified monoclonal antibody to Fas, induces apoptosis in human melanoma cells in vitro and delays the growth of melanoma xenotransplants. Oncol Rep 15: 409-415. |
| [190] |
Fellenberg J, Mau H, Scheuerpflug C, et al. (1997) Modulation of resistance to anti-APO-1-induced apoptosis in osteosarcoma cells by cytokines. Int J Cancer 72: 536-542. doi: 10.1002/(SICI)1097-0215(19970729)72:3<536::AID-IJC25>3.0.CO;2-8
|
| [191] |
Li Z, Xu Q, Peng H, et al. (2011) IFN-γ enhances HOS and U2OS cell lines susceptibility to γδ T cell-mediated killing through the Fas/Fas ligand pathway. Int Immnonopharmacol 11: 496-503. doi: 10.1016/j.intimp.2011.01.001
|
| [192] |
Garbán HJ, Bonavida B (1999) Nitric oxide sensitizes ovarian tumor cells to Fas-induced apoptosis. Gynecol Oncol 73: 257-264. doi: 10.1006/gyno.1999.5374
|
| [193] |
Jones NL, Day AS, Jennings HA, et al. (1999) Helicobacter pylori induces gastric epithelial cell apoptosis in association with increased Fas receptor expression. Infect Immun 67: 4237-4242. doi: 10.1128/IAI.67.8.4237-4242.1999
|
| [194] |
Wang J, Fan X, Lindholm C, et al. (2000) Helicobacter pylori modulates lymphoepithelial cell interactions leading to epithelial cell damage through Fas/Fas ligand interactions. Infect Immun 68: 4303-4311. doi: 10.1128/IAI.68.7.4303-4311.2000
|
| [195] |
Shin EC, Shin WC, Choi Y, et al. (2001) Effect of interferon-γ on the susceptibility to Fas (CD95/APO-1)-mediated cell death in human hepatoma cells. Cancer Immunol Immunother 50: 23-30. doi: 10.1007/s002620000166
|
| [196] |
Ahn EY, Pan G, Vickers SM, et al. (2002) IFN-γ upregulates apoptosis-related molecules and enhances Fas-mediated apoptosis in human cholangiocarcinoma. Int J Cancer 100: 445-451. doi: 10.1002/ijc.10516
|
| [197] |
Tsuji S, Hosotani R, Yonehara S, et al. (2003) Endogenous decoy receptor 3 blocks the growth inhibition signals mediated by Fas ligand in human pancreatic adenocarcinoma. Int J Cancer 106: 17-25. doi: 10.1002/ijc.11170
|
| [198] |
Selleck WA, Canfield SE, Hassen WA, et al. (2003) IFN-γ sensitization of prostate cancer cells to Fas-mediated death: a gene therapy approach. Mol Ther 7: 185-192. doi: 10.1016/S1525-0016(02)00040-0
|
| [199] |
Amrani A, Verdaguer J, Thiessen S, et al. (2000) IL-1α, IL-1β, and IFN-γ mark β cells for Fas-dependent destruction by diabetogenic CD4+ T lymphocytes. J Clin Invest 105: 459-468. doi: 10.1172/JCI8185
|
| [200] |
Mollah ZUA, Wail J, McKenzie MD, et al. (2011) The pro-apoptotic BH3-only protein Bid is dispensable for development of insulitis and diabetes in the non-obese diabetic mouse. Apoptosis 16: 822-830. doi: 10.1007/s10495-011-0615-z
|
| [201] |
Augstein P, Bahr J, Wachlin G, et al. (2004) Cytokines activate caspase-3 in insulinoma cells of diabetes-prone NOD mice directly and via upregulation of Fas. J Autoimmun 23: 301-309. doi: 10.1016/j.jaut.2004.09.006
|
| [202] |
Igoillo-Esteve M, Gurzov EN, Eizirik DL, et al. (2011) The transcription factor B-cell lymphoma (BCL)-6 modulates pancreatic β-cell inflammatory responses. Endocrinology 152: 447-456. doi: 10.1210/en.2010-0790
|
| [203] |
Allagnat F, Fukaya M, Nogueira TC, et al. (2012) C/EBP homologous protein contributes to cytokine-induced pro-inflammatory responses and apoptosis in β-cells. Cell Death Differ 19: 1836-1846. doi: 10.1038/cdd.2012.67
|
| [204] |
Nardelli TR, Vanzela EC, Benedicto KC, et al. (2018) Prolactin protects against cytokine-induced beta-cell death by NFκB and JNK inhibition. J Mol Endocrinol 61: 25-36. doi: 10.1530/JME-16-0257
|
| [205] |
Kawakami A, Eguchi K, Matsuoka N, et al. (1997) Modulation of Fas-mediated apoptosis of human thyroid epithelial cells by IgG from patients with Graves' disease (GD) and idiopathic myxoedema. Clin Exp Immunol 110: 434-439. doi: 10.1046/j.1365-2249.1997.4301447.x
|
| [206] |
Stassi G, Di Liberto D, Todaro M, et al. (2000) Control of target cell survival in thyroid autoimmunity by T helper cytokines via regulation of apoptotic proteins. Nat Immunol 1: 483-488. doi: 10.1038/82725
|
| [207] |
Pouly S, Becher B, Blain M, et al. (2000) Interferon-γ modulates human oligodendrocyte susceptibility to Fas-mediated apoptosis. J Neuropathol Exp Neurol 59: 280-286. doi: 10.1093/jnen/59.4.280
|
| [208] |
Yao Y, Lu S, Li H, et al. (2012) Low doses of exogenous interferon-γ attenuated airway inflammation through enhancing Fas/FasL-induced CD4+ T cell apoptosis in a mouse asthma model. J Interferon Cytokine Res 32: 534-541. doi: 10.1089/jir.2012.0016
|
| [209] |
Hallam DM, Capps NL, Travelstead AL, et al. (2000) Evidence for an interferon-related inflammatory reaction in the trisomy 16 mouse brain leading to caspase-1-mediated neuronal apoptosis. J Neuroimmunol 110: 66-75. doi: 10.1016/S0165-5728(00)00289-7
|
| [210] |
Ping L, Ogawa N, Sugai S (2005) Novel role of CD40 in Fas-dependent apoptosis of cultured salivary epithelial cells from patients with Sjögren's syndrome. Arthritis Rheum 52: 573-581. doi: 10.1002/art.20789
|
| [211] |
Fischer-Posovszky P, Hebestreit H, Hofmann AK, et al. (2006) Role of CD95-mediated adipocyte loss in autoimmune lipodystrophy. J Clin Endocrinol Metab 91: 1129-1135. doi: 10.1210/jc.2005-0737
|
| [212] |
Contassot E, Kerl K, Roques S, et al. (2008) Resistance to FasL and tumor necrosis factor-related apoptosis-inducing ligand-mediated apoptosis in Sézary syndrome T-cells associated with impaired death receptor and FLICE-inhibitory protein expression. Blood 111: 4780-4787. doi: 10.1182/blood-2007-08-109074
|
| [213] |
Conceiҫão-Silva F, Hahne M, Schröter M, et al. (1998) The resolution of lesions induced by Leishmania major in mice requires a functional Fas (APO-1, CD95) pathway of cytotoxicity. Eur J Immunol 28: 237-245. doi: 10.1002/(SICI)1521-4141(199801)28:01<237::AID-IMMU237>3.0.CO;2-O
|
| [214] |
Chakour R, Guler R, Bugnon M, et al. (2003) Both the Fas ligand and inducible nitric oxide synthase are needed for control of parasite replication within lesions in mice infected with Leishmania major whereas the contribution of tumor necrosis factor is minimal. Infect Immun 71: 5287-5295. doi: 10.1128/IAI.71.9.5287-5295.2003
|
| [215] |
Rosner D, Stoneman V, Littlewood T, et al. (2006) Interferon-γ induces Fas trafficking and sensitization to apoptosis in vascular smooth muscle cells via a PI3K- and Akt-dependent mechanism. Am J Pathol 168: 2054-2063. doi: 10.2353/ajpath.2006.050473
|
| [216] |
Esser P, Heimann K, Abts H, et al. (1995) CD95 (Fas/APO-1) antibody-mediated apoptosis of human retinal pigment epithelial cells. Biochem Biophys Res Comm 213: 1026-1034. doi: 10.1006/bbrc.1995.2231
|
| [217] |
Yang Z, Gagarin D, St. Laurent III G, et al. (2009) Cardiovascular inflammation and lesion cell apoptosis. A novel connection via the interferon-inducible immunoproteasome. Arterioscler Thromb Vasc Biol 29: 1213-1219. doi: 10.1161/ATVBAHA.109.189407
|
| [218] |
Zhaorigetu S, Yang Z, Toma I, et al. (2011) Apolipoprotein L6, induced in atherosclerotic lesions promotes apoptosis and blocks Beclin 1-dependent autophagy in atherosclerotic cells. J Biol Chem 286: 27389-27398. doi: 10.1074/jbc.M110.210245
|
| [219] |
Catalan MP, Subirá D, Reyero A, et al. (2003) Regulation of apoptosis by lethal cytokines in human mesothelial cells. Kidney Int 64: 321-330. doi: 10.1046/j.1523-1755.2003.00062.x
|
| [220] |
Wang XY, Crowston JG, Zoellner H, et al. (2007) Interferon-α and interferon-γ sensitize human tenon fibroblasts to mitomycin-C. Invest Ophthalmol Vis Sci 48: 3655-3661. doi: 10.1167/iovs.06-1121
|
| [221] |
Zhang X, Chen W, De Paiva CS, et al. (2011) Interferon-γ exacerbates dry eye-induced apoptosis in conjunctiva through dual apoptotic pathways. Invest Ophthalmol Vis Sci 52: 6279-6285. doi: 10.1167/iovs.10-7081
|
| [222] |
Nagafuji K, Shibuya T, Harada M, et al. (1995) Functional expression of Fas antigen (CD95) on hematopoietic progenitor cells. Blood 86: 883-889. doi: 10.1182/blood.V86.3.883.883
|
| [223] |
Oyaizu N, Mc Closkey TW, Than S, et al. (1996) Inhibition of CD4 cross-linking-induced lymphocytes apoptosis by vesnarinone as a novel immunomodulating agent: vesnarinone inhibits Fas expression and apoptosis by blocking cytokine secretion. Blood 87: 2361-2368. doi: 10.1182/blood.V87.6.2361.bloodjournal8762361
|
| [224] |
Sato T, Selleri C, Anderson S, et al. (1997) Expression and modulation of cellular receptors for interferon-γ, tumour necrosis factor, and Fas on human bone marrow CD34+ cells. Br J Haematol 97: 356-365. doi: 10.1046/j.1365-2141.1997.562704.x
|
| [225] |
Schneider E, Moreau G, Arnould A, et al. (1999) Increased fatal and extramedullary hematopoiesis in Fas-deficient C57BL/6-lpr/lpr mice. Blood 94: 2613-2621. doi: 10.1182/blood.V94.8.2613.420k33_2613_2621
|
| [226] |
Erie AJ, Samsel L, Takaku T, et al. (2011) MHC class II upregulation and colocalization with Fas in experimental models of immune-mediated bone marrow failure. Exp Hematol 39: 837-849. doi: 10.1016/j.exphem.2011.05.005
|
| [227] |
Kohara H, Kitaura H, Fujimura Y, et al. (2011) IFN-γ directly inhibits TNF-α-induced osteoclastgenesis in vitro and in vivo and induces apoptosis mediated by Fas/Fas ligand interactions. Immunol Lett 137: 53-61. doi: 10.1016/j.imlet.2011.02.017
|
| [228] |
Liu Y, Wang L, Kikuiri T, et al. (2011) Mesenchymal stem cell-based tissue regeneration is governed by recipient T lymphocytes via IFN-γ and TNF-α. Nat Med 17: 1594-1601. doi: 10.1038/nm.2542
|
| [229] |
Chen J, Feng X, Desierto MJ, et al. (2015) IFN-γ-mediated hematopoietic cell destruction in murine models of immune-mediated bone marrow failure. Blood 126: 2621-2631. doi: 10.1182/blood-2015-06-652453
|
| [230] |
Xu J, Wang Y, Li J, et al. (2016) IL-12p40 impairs mesenchymal stem cell-mediated bone regeneration via CD4+ T cells. Cell Death Differ 23: 1941-1951. doi: 10.1038/cdd.2016.72
|
| [231] |
Li X, Shang B, Li YN (2019) IFNγ and TNFα synergistically induce apoptosis of mesenchymal stem/stromal cells via induction of nitric oxide. Stem Cell Res Ther. 10: 18. doi: 10.1186/s13287-018-1102-z
|
| [232] |
Luttmann W, Opfer A, Dauer E, et al. (1998) Differential regulation of CD95 (Fas/APO-1) expression in human blood eosinophils. Eur J Immunol 28: 2057-2065. doi: 10.1002/(SICI)1521-4141(199807)28:07<2057::AID-IMMU2057>3.0.CO;2-T
|
| [233] |
Luttmann W, Dauer E, Schmidt S, et al. (2000) Effects of interferon-γ and tumour necrosis factor-α on CD95/Fas ligand-mediated apoptosis in human blood eosinophils. Scand J Immunol 51: 54-59. doi: 10.1046/j.1365-3083.2000.00645.x
|
| [234] |
Zizzo G, Cohen PL (2013) IL-17 stimulates differentiation of human anti-inflammatory macrophages and phagocytosis of apoptotic neutrophils in response to IL-10 and glucocorticoids. J Immunol 190: 5237-5246. doi: 10.4049/jimmunol.1203017
|
| [235] |
Hagmann BR, Odermatt A, Kaufmann T, et al. (2016) Balance between IL-3 and type I interferons and their interrelationship with FasL dictates lifespan and effector functions of human basophils. Clin Exp Allergy 47: 71-84. doi: 10.1111/cea.12850
|
| [236] |
Chung IJ, Dai C, Krantz SB (2003) Stem cell factor increases the expression of FLIP that inhibits IFN-γ-induced apoptosis in human erythroid progenitor cells. Blood 101: 1324-1328. doi: 10.1182/blood-2002-06-1720
|
| [237] |
Müschen M, Warskulat U, Peters-Regehr T, et al. (1999) Involvement of CD95 (Apo-1/Fas) ligand expressed by rat Kupffer cells in hepatic immunoregulation. Gastroenterology 116: 666-677. doi: 10.1016/S0016-5085(99)70189-7
|
| [238] |
Bárcena A, Park SW, Banapour B, et al. (1996) Expression of Fas/CD95 and Bcl-2 by primitive hematopoietic progenitors freshly isolated from human fetal liver. Blood 88: 2013-2025. doi: 10.1182/blood.V88.6.2013.bloodjournal8862013
|
| [239] |
Oh JE, Shim KY, Lee JI, et al. (2017) 1-Methyl-L-tryptophan promotes the apoptosis of hepatic stellate cells arrested by interferon-γ by increasing the expression of IFN-γRβ, IRF-1 and Fas. Int J Mol Med 40: 576-582. doi: 10.3892/ijmm.2017.3043
|
| [240] |
He W, Yang C, Xia L, et al. (2014) CD4+ T cells from food allergy model are resistant to TCR-dependent apoptotic induction. Cytokine 68: 32-39. doi: 10.1016/j.cyto.2014.03.010
|
| [241] |
Lightfoot YL, Chen J, Mathews CE (2011) Role of the mitochondria in immune-mediated apoptotic death of the human pancreatic β cell line βLox5. Plos One 6: e20617. doi: 10.1371/journal.pone.0020617
|
| [242] | Sun Q, Xiang RL, Yang YL, et al. (2013) Suppressor of cytokine signaling 1 protects rat pancreatic islets from cytokine-induced apoptosis through Janus kinase/signal transducers and activators of transcription pathway. Chin Med J 126: 4048-4053. |
| [243] |
Quirk SM, Cowan RG, Huber SH (1997) Fas antigen-mediated apoptosis of ovarian surface epithelial cells. Endocrinology 138: 4558-4566. doi: 10.1210/endo.138.11.5508
|
| [244] |
Taniguchi H, Yokomizo Y, Okuda K (2002) Fas-Fas ligand system mediates luteal cell death in bovine corpus luteum. Biol Reprod 66: 754-759. doi: 10.1095/biolreprod66.3.754
|
| [245] |
Galvao AM, Ramilo DW, Skarzynski DJ, et al. (2010) Is Fas/Fas ligand system involved in equine corpus luteum functional regression? Biol Reprod 83: 901-908. doi: 10.1095/biolreprod.110.084699
|
| [246] |
Hojo T, Al-zi'abi O, Komiyama J, et al. (2010) Expression and localization of FLIP, an anti-apoptotic factor, in the bovine corpus luteum. J Reprod Dev 56: 230-235. doi: 10.1262/jrd.09-185S
|
| [247] |
Woclawek-Potocka I, Kowalczyk-Zieba I, Tylingo M, et al. (2013) Effects of lysophosphatidic acid on tumor necrosis factor α and interferon γ action in the bovine corpus luteum. Mol Cell Endrocrinol 377: 103-111. doi: 10.1016/j.mce.2013.07.005
|
| [248] |
Quirk SM, Porter DA, Huber SC, et al. (1998) Potentiation of Fas-mediated apoptosis of murine granulosa cells by interferon-γ, tumor necrosis factor-α, and cycloheximide. Endocrinology 139: 4860-4869. doi: 10.1210/endo.139.12.6353
|
| [249] |
Lee HJ, Kim JY, Park JE, et al. (2016) Induction of Fas-mediated apoptosis by interferon-γ is dependent on granulosa cell differentiation and follicular maturation in the rat ovary. Dev Reprod 20: 315-329. doi: 10.12717/DR.2016.20.4.315
|
| [250] |
Aschkenazi S, Straszewski S, Verwer KMA, et al. (2002) Differential regulation and function of the Fas/Fas ligand system in human trophoblast cells. Biol Reprod 66: 1853-1861. doi: 10.1095/biolreprod66.6.1853
|
| [251] |
Balkundi DR, Ziegler JA, Watchko JF, et al. (2003) Regulation of FasL/Fas in human trophoblasts: possible implications for chorioamnionitis. Biol Reprod 69: 718-724. doi: 10.1095/biolreprod.102.013102
|
| [252] |
Riccioli A, Starace D, D'Alessio A, et al. (2000) TNF-α and IFN-γ regulate expression and function of the Fas system in the seminiferous epithelium. J Immunol 165: 743-749. doi: 10.4049/jimmunol.165.2.743
|
| [253] |
González-Cuadrado S, López-Armada MJ, Gómez-Guerrero C, et al. (1996) Anti-Fas antibodies induce cytolysis and apoptosis in cultured human mesangial cells. Kidney Int 49: 1064-1070. doi: 10.1038/ki.1996.155
|
| [254] |
Tsukinoki T, Sugiyama H, Sunami R, et al. (2004) Mesangial cell Fas ligand: upregulation in human lupus nephritis and NF-κB-mediated expression in cultured human mesangial cells. Clin Exp Nephrol 8: 196-205. doi: 10.1007/s10157-004-0301-3
|
| [255] | Lorz C, Ortiz A, Justo P, et al. (2000) Proapoptotic Fas ligand is expressed by normal kidney tubular epithelium and injured glomeruli. J Am Soc Nephrol 11: 1266-1277. |
| [256] |
Spanaus KS, Schlapbach R, Fontana A (1998) TNF-α and INF-γ render microglia sensitive to Fas ligand-induced apoptosis by induction of Fas expression and down-regulation of Bcl-2 and Bcl-xL. Eur J Immunol 28: 4398-4408. doi: 10.1002/(SICI)1521-4141(199812)28:12<4398::AID-IMMU4398>3.0.CO;2-Y
|
| [257] |
Schlapbach R, Spanaus KS, Malipiero U, et al. (2000) TGF-β induces the expression of the FLICE-inhibitory protein and inhibits Fas-mediated apoptosis of microglia. Eur J Immunol 30: 3680-3688. doi: 10.1002/1521-4141(200012)30:12<3680::AID-IMMU3680>3.0.CO;2-L
|
| [258] |
Falsig J, Latta M, Leist M (2004) Defined inflammatory states in astrocyte cultures: correlation with susceptibility towards CD95-driven apoptosis. J Neurochem 88: 181-193. doi: 10.1111/j.1471-4159.2004.02144.x
|
| [259] |
Coque E, Salsac C, Espinosa-Carrasco G, et al. (2019) Cytotoxic CD8+ T lymphocytes expressing ALS-causing SOD1 mutant selectively trigger death of spinal motoneurons. Proc Natl Acad Sci USA 116: 2312-2317. doi: 10.1073/pnas.1815961116
|
| [260] |
Sayama K, Yonehara S, Watanabe Y, et al. (1994) Expression of Fas antigen on keratinocytes in vivo and induction of apoptosis in cultured keratinocytes. J Invest Dermatol 103: 330-334. doi: 10.1111/1523-1747.ep12394858
|
| [261] |
Viard-Leveugle I, Gaide O, Jankovic D, et al. (2013) TNF-α and IFN-γ are potential inducers of Fas-mediated keratinocyte apoptosis through activation of inducible nitric oxide synthase in toxic epidermal necrolysis. J Invest Dermatol 133: 489-498. doi: 10.1038/jid.2012.330
|
| [262] |
Gao Z, Jin YQ, Wu W (2017) SOCS3 treatment prevents the development of alopecia areata by inhibiting CD8+ T cell-mediated autoimmune destruction. Oncotarget 8: 33432-33443. doi: 10.18632/oncotarget.16504
|
| [263] |
Li JH, Kluger MS, Madge LA, et al. (2002) Interferon-γ augments CD95 (APO-1/Fas) and pro-caspase-8 expression and sensitizes human vascular endothelial cells to CD95-mediated apoptosis. Am J Pathol 161: 1485-1495. doi: 10.1016/S0002-9440(10)64424-0
|
| [264] |
Yamaoka-Tojo M, Yamaguchi S, Nitobe J, et al. (2003) Dual response to Fas ligation in human endothelial cells: apoptosis and induction of chemokines, interleukin-8 and monocyte chemoattractant protein-1. Coron Artery Dis 14: 89-94. doi: 10.1097/00019501-200302000-00010
|
| [265] |
Shigeta A, Tada Y, Wang JY, et al. (2012) CD40 amplifies Fas-mediated apoptosis: a mechanism contributing to emphysema. Am J Physiol Lung Cell Mol Physiol 303: L141-L151. doi: 10.1152/ajplung.00337.2011
|
| [266] |
Kawakami A, Eguchi K, Matsuoka N, et al. (1996) Thyroid-stimulating hormone inhibits Fas antigen mediated apoptosis of human thyrocytes in vitro. Endocrinology 137: 3163-3169. doi: 10.1210/endo.137.8.8754734
|
| [267] |
Mezosi E, Yamazaki H, Bretz JD, et al. (2002) Aberrant apoptosis in thyroid epithelial cells from goiter nodules. J Clin Endocrinol Metab 87: 4264-4272. doi: 10.1210/jc.2002-020111
|
| [268] |
Mezosi E, Wang SH, Utsugi S, et al. (2005) Induction and regulation of Fas-mediated apoptosis in human thyroid epithelial cells. Mol Endocrinol 19: 804-811. doi: 10.1210/me.2004-0286
|
| [269] |
Wang SH, van Antwerp M, Kuick R, et al. (2007) Microarray analysis of cytokine activation of apoptosis pathways in the thyroid. Endocrinology 148: 4844-4852. doi: 10.1210/en.2007-0126
|
| [270] |
Fang Y, Braley-Mullen H (2008) Cultured murine thyroid epithelial cells expressing transgenic Fas-associated death-like interleukin-1β converting enzyme inhibitory protein are protected from Fas-mediated apoptosis. Endocrinology 149: 3321-3329. doi: 10.1210/en.2008-0080
|
| [271] | Marsumura R, Umemiya K, Goto T, et al. (2000) Interferon gamma and tumor necrosis factor alpha induce Fas expression and anti-Fas mediated apoptosis in a salivary ductal cell line. Clin Exp Rheumatol 18: 311-318. |
| [272] |
Abu-Helu RF, Dimitriou ID, Kapsogeorgou EK, et al. (2001) Induction of salivary gland epithelial cell injury in Sjogren's syndrome: in vitro assessment of T cell-derived cytokines and Fas protein expression. J Autoimmun 17: 141-153. doi: 10.1006/jaut.2001.0524
|
| [273] |
Ruemmele FM, Russo P, Beaulieu JF, et al. (1999) Susceptibility to Fas-induced apoptosis in human nontumoral enterocytes: role of costimulatory factors. J Cell Physiol 181: 45-54. doi: 10.1002/(SICI)1097-4652(199910)181:1<45::AID-JCP5>3.0.CO;2-Q
|
| [274] |
Bharhani MS, Borojevic R, Basak S, et al. (2006) IL-10 protects mouse intestinal epithelial cells from Fas-induced apoptosis via modulating Fas expression and altering caspase-8 and FLIP expression. Am J Physiol Gastrointest Liver Physiol 291: G820-G829. doi: 10.1152/ajpgi.00438.2005
|
| [275] | De Saint Jean M, Debbasch C, Rahmani M, et al. (2000) Fas-and interferon γ-induced apoptosis in Chang conjunctival cells: further investigations. Invest Ophthalomol Vis Sci 41: 2531-2543. |
| [276] |
Gao J, Sana R, Calder V, et al. (2013) Mitochondrial permeability transition pore in inflammatory apoptosis of human conjunctival epithelial cells and T cells: effect of cyclosporine A. Invest Ophthalmol Vis Sci 54: 4717-4733. doi: 10.1167/iovs.13-11681
|
| [277] |
Nakamura M, Matute-Bello G, Liles WC, et al. (2004) Differential response of human lung epithelial cells to Fas-induced apoptosis. Am J Pathol 164: 1949-1958. doi: 10.1016/S0002-9440(10)63755-8
|
| [278] |
Arai M, Yoshioka S, Nishimura R, et al. (2014) Fas/FasL-mediated cell death in the bovine endometrium. Anim Reprod Sci. 151: 97-104. doi: 10.1016/j.anireprosci.2014.10.004
|
| [279] |
Frankel SK, Cosgrove GP, Cha SI, et al. (2006) TNF-α sensitizes normal and fibrotic human lung fibroblasts to Fas-induced apoptosis. Am J Respir Cell Mol Biol 34: 293-304. doi: 10.1165/rcmb.2005-0155OC
|
| [280] |
Lee JW, Oh JE, Rhee KJ, et al. (2019) Co-treatment with interferon-γ and 1-methyl tryptophan ameliorates cardiac fibrosis through cardiac myofibroblasts apoptosis. Mol Cell Biochem 458: 197-205. doi: 10.1007/s11010-019-03542-7
|
| [281] |
Fluhr H, Krenzer S, Stein GM, et al. (2007) Interferon-γ and tumor necrosis factor-α sensitize primarily resistant human endometrial stromal cells to Fas-mediated apoptosis. J Cell Sci 120: 4126-4133. doi: 10.1242/jcs.009761
|
| [282] |
Boeddeker SJ, Baston-Buest DM, Fehm T, et al. (2015) Decidualization and syndecan-1 knock down sensitize endometrial stromal cells to apoptosis induced by embryonic stimuli. Plos One 10: e0121103. doi: 10.1371/journal.pone.0121103
|
| [283] |
Yamada K, Takane-Gyotoku N, Yuan X, et al. (1996) Mouse islet cell lysis mediated by interleukin-1-induced Fas. Diabetologia 39: 1306-1312. doi: 10.1007/s001250050574
|
| [284] |
Roth W, Wagenknecht B, Dichgans J, et al. (1998) Interferon-α enhances CD95L-induced apoptosis of human malignant glioma cells. J Neuroimmunol 87: 121-129. doi: 10.1016/S0165-5728(98)00079-4
|
| [285] |
Dey BR, Yang YG, Szot GL, et al. (1998) Interleukin-12 inhibits Graft-Versus-Host disease through a Fas-mediated mechanism associated with alterations in donor T-cell activation and expansion. Blood 91: 3315-3322. doi: 10.1182/blood.V91.9.3315
|
| [286] |
Kobayashi T, Okamoto K, Kobata T, et al. (1999) Tumor necrosis factor α regulation of the Fas-mediated apoptosis-signaling pathway in synovial cells. Arthritis Rheum 42: 519-526. doi: 10.1002/1529-0131(199904)42:3<519::AID-ANR17>3.0.CO;2-Q
|
| [287] |
Houghton J, Macera-Bloch LS, Harrison L, et al. (2000) Tumor necrosis factor alpha and interleukin 1β up-regulate gastric mucosal Fas antigen expression in Helicobacter pylori infection. Infect Immun 68: 1189-1195. doi: 10.1128/IAI.68.3.1189-1195.2000
|
| [288] |
Kimura K, Gelmann EP (2000) Tumor necrosis factor-α and Fas activate complementary Fas-associated death domain-dependent pathways that enhance apoptosis induced by γ-irradiation. J Biol Chem 275: 8610-8617. doi: 10.1074/jbc.275.12.8610
|
| [289] |
Reddy P, Teshima T, Kukuruga M, et al. (2001) Interleukin-18 regulates acute graft-versus-host disease by enhancing Fas-mediated donor T cell apoptosis. J Exp Med 10: 1433-1440. doi: 10.1084/jem.194.10.1433
|
| [290] |
Elzey BD, Griffith TS, Herndon JM, et al. (2001) Regulation of Fas ligand-induced apoptosis by TNF. J Immunol 167: 3049-3056. doi: 10.4049/jimmunol.167.6.3049
|
| [291] |
Sharief MK, Semra YK (2002) Down-regulation of survivin expression in T lymphocytes after interferon beta-1a treatment in patients with multiple sclerosis. Arch Neurol 59: 1115-1121. doi: 10.1001/archneur.59.7.1115
|
| [292] |
Park SM, Park HY, Lee TH (2003) Functional effects of TNF-α on a human follicular dendritic cell line: persistent NF-κB activation and sensitization for Fas-mediated apoptosis. J Immunol 171: 3955-3962. doi: 10.4049/jimmunol.171.8.3955
|
| [293] |
Schlosser SF, Schuler M, Christoph PB, et al. (2003) Ribavirin and alpha interferon enhance death receptor-mediated apoptosis and caspase activation in human hepatoma cells. Antimicrob Agents Chemother 47: 1912-1921. doi: 10.1128/AAC.47.6.1912-1921.2003
|
| [294] |
Schneider E, Tonanny MB, Lisbonne M, et al. (2004) Pro-Th1 cytokines promote Fas-dependent apoptosis of immature peripheral basophils. J Immnunol 172: 5262-5268. doi: 10.4049/jimmunol.172.9.5262
|
| [295] |
Dondi E, Roué G, Yuste VJ (2004) A dual role of IFN-α in the balance between proliferation and death of human CD4+ T lymphocytes during primary response. J Immunol 173: 3740-3747. doi: 10.4049/jimmunol.173.6.3740
|
| [296] |
Kelly JD, Dai J, Eschwege P, et al. (2004) Downregulation of Bcl-2 sensitizes interferon-resistant renal cancer cells to Fas. Br J Cancer 91: 164-170. doi: 10.1038/sj.bjc.6601895
|
| [297] |
Wu X, Pan G, McKenna MA, et al. (2005) RANKL regulates Fas expression and Fas-mediated apoptosis in osteoclasts. J Bone Mineral Res 20: 107-116. doi: 10.1359/JBMR.041022
|
| [298] |
Park SM, Kim S, Choi JS, et al. (2005) TGF-β inhibits Fas-mediated apoptosis of a follicular dendritic cell line by down-regulating the expression of Fas and caspase-8: counteracting role of TGF-β on TNF sensitization of Fas-mediated apoptosis. J Immunol 174: 6169-6175. doi: 10.4049/jimmunol.174.10.6169
|
| [299] |
Drynda A, Quax PHA, Neumann M, et al. (2005) Gene transfer of tissue inhibitor of metalloproteinases-3 reverses the inhibitory effects of TNF-α on Fas-induced apoptosis in rheumatoid arthritis synovial fibroblasts. J Immunol 174: 6524-6531. doi: 10.4049/jimmunol.174.10.6524
|
| [300] |
Lindkvist A, Ivarsson K, Jernberg-Wiklund H, et al. (2006) Interferon-induced sensitization to apoptosis is associated with repressed transcriptional activity of the hTERT promoter in multiple myeloma. Biochem Biophys Res Comm 341: 1141-1148. doi: 10.1016/j.bbrc.2006.01.068
|
| [301] |
Huerta-Yepez S, Vega M, Garban H, et al. (2006) Involvement of the TNF-α autocrine-paracrine loop, via NF-κB and YY1, in the regulation of tumor cell resistance to Fas-induced apoptosis. Clin Immunol 120: 297-309. doi: 10.1016/j.clim.2006.03.015
|
| [302] |
Corazza N, Jakob S, Schaer C, et al. (2006) TRAIL receptor-mediated JNK activation and Bim phosphorylation critically regulate Fas-mediated liver damage and lethality. J Clin Invest 116: 2493-2499. doi: 10.1172/JCI27726
|
| [303] |
Yang J, Epling-Burnette PK, Painter JS, et al. (2008) Antigen activation and impaired Fas-induced death-inducing signaling complex formation in T-large-granular lymphocyte leukemia. Blood 111: 1610-1616. doi: 10.1182/blood-2007-06-093823
|
| [304] |
Nihal M, Ahsan H, Siddiqui IA, et al. (2009) (-)-Epigallocatechin-3-gallate (EGCG) sensitizes melanoma cells to interferon induced growth inhibition in a mouse model of human melanoma. Cell Cycle 8: 2057-2063. doi: 10.4161/cc.8.13.8862
|
| [305] |
Wu J, Wood GS (2011) Reduction of Fas/CD95 promoter methylation, upregulation of Fas protein, and enhancement of sensitivity to apoptosis in cutaneous T-cell lymphoma. Arch Dermatol 147: 443-449. doi: 10.1001/archdermatol.2010.376
|
| [306] |
Schmich K, Schlatter R, Corazza N, et al. (2011) Tumor necrosis factor α sensitizes primary murine hepatocytes to Fas/CD95-induced apoptosis in a Bim- and Bid-dependent manner. Hepatology 53: 282-292. doi: 10.1002/hep.23987
|
| [307] |
Roos WP, Jöst E, Belohlavek C, et al. (2011) Intrinsic anticancer drug resistance of malignant melanoma cells is abrogated by IFN-β and valproic acid. Cancer Res 71: 4150-4160. doi: 10.1158/0008-5472.CAN-10-3498
|
| [308] |
Fraietta JA, Mueller YM, Yang G, et al. (2013) Type I interferon upregulates Bak and contributes to T cell loss during human immunodeficiency virus (HIV) infection. Plos Pathog 9: e1003658. doi: 10.1371/journal.ppat.1003658
|
| [309] |
Lutz A, Sanwald J, Thomas M, et al. (2014) Interleukin-1β enhances FasL-induced caspase-3/-7 activity without increasing apoptosis in primary mouse hepatocytes. Plos One 9: e115603. doi: 10.1371/journal.pone.0115603
|
| [310] |
Faletti L, Peintner L, Neumann S, et al. (2018) TNFα sensitizes hepatocytes to FasL-induced apoptosis by NFκB-mediated Fas upregulation. Cell Death Dis 9: 909. doi: 10.1038/s41419-018-0935-9
|
| [311] |
Aggarwal BB, Gupta SC, Kim JH (2012) Historical perspectives on tumor necrosis factor and its superfamily: 25 years later, a golden journey. Blood 119: 651-655. doi: 10.1182/blood-2011-04-325225
|
| [312] |
van Boxel-Dezaire AHH, Rani MRS, Stark GR (2006) Complex modulation of cell type-specific signaling in response to type I interferons. Immunity 25: 361-372. doi: 10.1016/j.immuni.2006.08.014
|
| [313] |
Hunter CA, Timans J, Pisacane P, et al. (1997) Comparison of the effects of interleukin-1α, interleukin-1β and interferon-γ-inducing factor on the production of interferon-γ by natural killer. Eur J Immunol 27: 2787-2792. doi: 10.1002/eji.1830271107
|
| [314] |
Bonta IL, Ben-Efraim S (1993) Involvement of inflammatory mediators in macrophage antitumor activity. J Leukoc Biol 54: 613-626. doi: 10.1002/jlb.54.6.613
|
| [315] |
Stylianou E, Saklatvala J (1998) Interleukin-1. Int J Biochem Cell Biol 30: 1075-1079. doi: 10.1016/S1357-2725(98)00081-8
|
| [316] |
Miwa K, Asano M, Horai R, et al. (1998) Caspase 1-indenendent IL-1β release and inflammation induced by the apoptosis inducer Fas ligand. Nat Med 4: 1287-1292. doi: 10.1038/3276
|
| [317] |
Linkermann A, Qian J, Lettau M, et al. (2005) Considering Fas ligand as a target for therapy. Expert Opin Ther Targets 9: 119-134. doi: 10.1517/14728222.9.1.119
|
| [318] |
Villa-Morales M, Fernández-Piqueras (2012) Targeting the Fas/FasL signaling pathway in cancer therapy. Expert Opin Ther Targets 16: 85-101. doi: 10.1517/14728222.2011.628937
|
| [319] |
Michael-Robinson JM, Pandeya N, Cummings MC, et al. (2003) Fas ligand and tumour counter-attack in colorectal cancer stratified according to microsatellite instability status. J Pathol 201: 46-54. doi: 10.1002/path.1406
|
| [320] |
Ogasawara J, Watanabe-Fukunaga R, Adachi M, et al. (1993) Lethal effect of the anti-Fas antibody in mice. Nature 26: 806-809. doi: 10.1038/364806a0
|
| [321] |
Tsujimoto Y, Shimizu S (2000) Bcl-2 family: life-or-death switch. FEBS Lett 466: 6-10. doi: 10.1016/S0014-5793(99)01761-5
|
| [322] |
Zhuang S, Dermirs JT, Kochevar IE (2001) Protein kinase C inhibits singlet oxygen-induced apoptosis by decreasing caspase-8 activation. Oncogene 20: 6764-6776. doi: 10.1038/sj.onc.1204867
|
| [323] |
Suzuki Y, Ono Y, Hirabayashi Y (1998) Rapid and specific reactive oxygene species generation during Fas-mediated apoptosis. FEBS Lett 425: 209-212. doi: 10.1016/S0014-5793(98)00228-2
|
| [324] |
Reinehr R, Becker S, Eberle A, et al. (2005) Involvement of NADPH oxidase isoforms and Src family kinases in CD95-dependent hepatocyte apoptosis. J Biol Chem 280: 27179-27194. doi: 10.1074/jbc.M414361200
|
| [325] |
Selleri C, Sato T, Raiola AM, et al. (1997) Induction of nitric oxide synthase is involved in the mechanism of Fas-mediated apoptosis in heamopoietic cells. Br J Heamatol 99: 481-489. doi: 10.1046/j.1365-2141.1996.4323240.x
|
| [326] | Solano-Gálvez SG, Abadi-Chiriti J, Gutiérrez-Velez L, et al. (2018) Apoptosis: activation and inhibition in health and disease. Med Sci 6: 54. |
| [327] |
Seyrek K, Lavik IN (2019) Modulation of CD95-mediated signaling by post-translational modifications: towards understanding CD95 signaling networks. Apoptosis 24: 385-394. doi: 10.1007/s10495-019-01540-0
|
| [328] |
Pitti RM, Marsters SA, Lawrence DA, et al. (1998) Genomic amplification of a decoy receptor for Fas ligand in lung and colon cancer. Nature 396: 699-703. doi: 10.1038/25387
|
| [329] |
Liu W, Ramagopal U, Cheng H, et al. (2016) Crystal structure of the complex of human FasL and its decoy receptor DcR3. Structure 24: 2016-2023. doi: 10.1016/j.str.2016.09.009
|
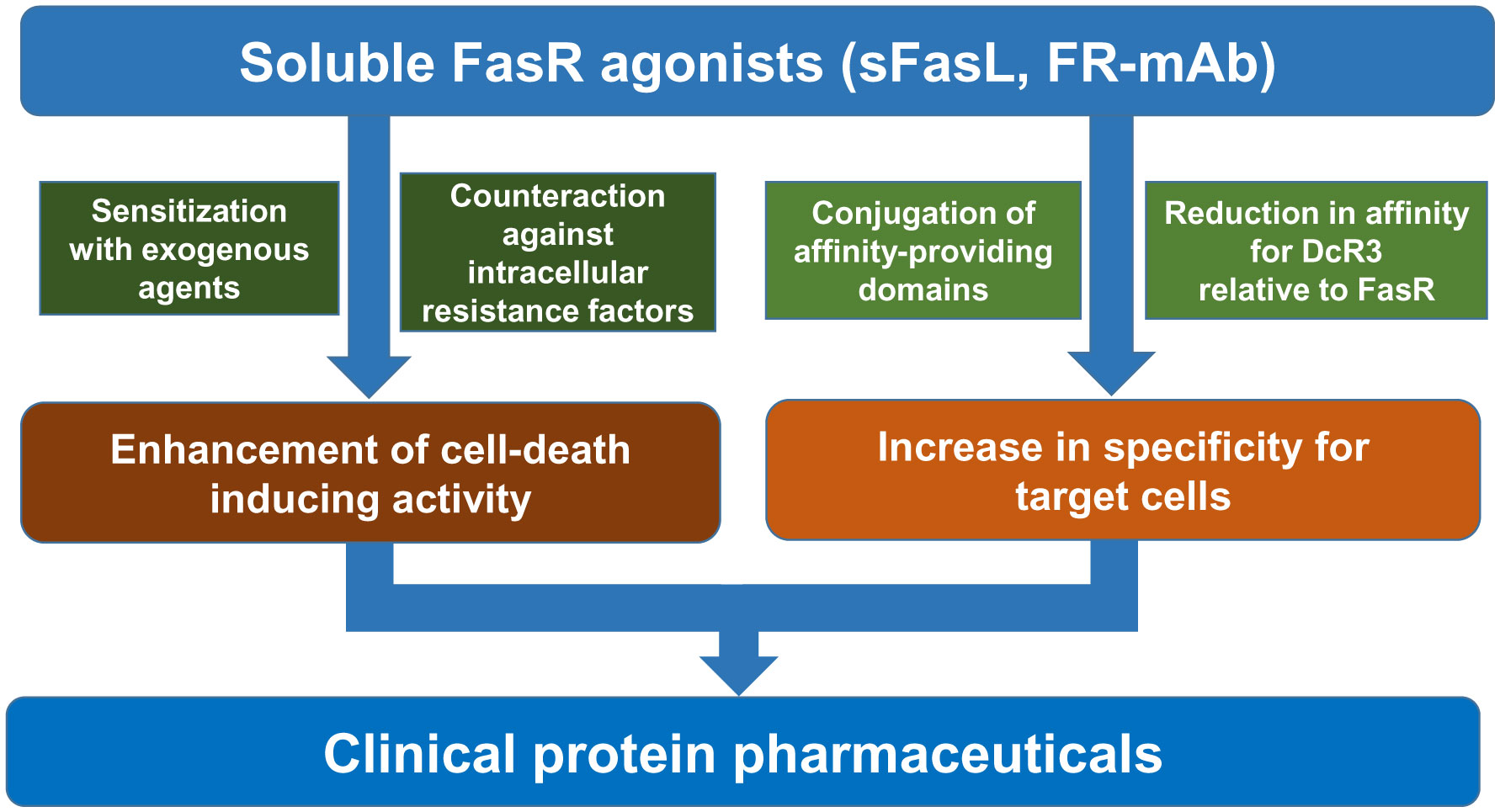
Figures(1) / Tables(3)

Michiro Muraki. Sensitization to cell death induced by soluble Fas ligand and agonistic antibodies with exogenous agents: A review[J]. AIMS Medical Science, 2020, 7(3): 122-203. doi: 10.3934/medsci.2020011







 DownLoad:
DownLoad: