Citation: Eugene Verkhovsky, Paul Naylor, Sindhuri Benjaram, Shanker Das Kundumadam, Murray Ehrinpreis, Milton Mutchnick. Racial disparity in chronic hepatitis B infection in a predominately African American urban clinic population[J]. AIMS Medical Science, 2018, 5(4): 378-385. doi: 10.3934/medsci.2018.4.378

| [1] |
Ioannou GN (2011) Hepatitis B virus in the United States: infection, exposure, and immunity rates in a nationally representative survey. Ann Intern Med 154: 319–328. doi: 10.7326/0003-4819-154-5-201103010-00006

|
| [2] | Yuen MF, Chen DS, Dusheiko GM, et al. (2018) Hepatitis B virus infection. Nat Rev Dis Primers 4: 2056–2676. |
| [3] |
Kowdley KV, Wang CC, Welch S, et al. (2012) Prevalence of chronic hepatitis B among foreign-born persons living in the United States by country of origin. Hepatology 56: 422–433. doi: 10.1002/hep.24804
|
| [4] |
McQuillan GM, Coleman PJ, Kruszon-Moran D, et al. (1999) Prevalence of hepatitis B virus infection in the United States: the National Health and Nutrition Examination Surveys, 1976 through 1994. Am J Public Health 89: 14–18. doi: 10.2105/AJPH.89.1.14
|
| [5] |
Ford KA, Tanapanpanit O, Reddy KR (2014) Hepatitis B and C in African Americans: Current status and continued challenges. Clin Gastroenterol Hepatol 12: 738–748. doi: 10.1016/j.cgh.2013.06.006
|
| [6] |
McQuillan GM, Kruszon-Moran D, Kottiri BJ, et al. (2004) Racial and ethnic differences in the seroprevalence of 6 infectious diseases in the United States: data from NHANES III, 1988-1994. Am J Public Health 94: 1952–1958. doi: 10.2105/AJPH.94.11.1952
|
| [7] | MacLachlan JH, Cowie BC (2015) Hepatitis B Virus Epidemiology. Cold Spring Harb Perspect Med 5: 1–10. |
| [8] |
Azmi AN, Tan SS, Mohamed R (2014) Practical approach in hepatitis B e antigen-negative individuals to identify treatment candidates. World J Gastroenterol 20: 12045–12055. doi: 10.3748/wjg.v20.i34.12045
|
| [9] | Widjaja D, Yarlagadda S, Singu BS, et al. (2007) Characteristics of patients with chronic hepatitis-B virus infection in an urban hospital. J Natl Med Assoc 99: 384–388. |
| [10] | Agarwal K, Brunetto M, Seto WK, et al. (2018) 96 weeks treatment of tenofovir alafenamide vs. tenofovir disoproxil fumarate for hepatitis B virus infection. J Hepatology 68: 672–681. |
| [11] |
Tang LSY, Covert E, Wilson E, et al. (2018) Chronic Hepatitis B Infection: A Review. JAMA 319: 1802–1813. doi: 10.1001/jama.2018.3795
|
| [12] |
Tawada A, Kanda T, Yokosuka O (2015) Current and future directions for treating hepatitis B virus infection. World J Hepatol 7: 1541–1552. doi: 10.4254/wjh.v7.i11.1541
|
| [13] |
Sundaram V, Kowdley K (2015) Management of chronic hepatitis B infection. BMJ 351: h4263. doi: 10.1136/bmj.h4263
|
| [14] |
Tang CM, Yau TO, Yu J (2014) Management of chronic hepatitis B infection: Current treatment guidelines, challenges, and new developments. World J. Gastroenterol 20: 6262–6278. doi: 10.3748/wjg.v20.i20.6262
|
| [15] |
Lok AS, McMahon BJ, Brown RS Jr, et al. (2016) Antiviral therapy for chronic hepatitis B viral infection in adults: A systematic review and meta-analysis. Hepatology 63: 284–306. doi: 10.1002/hep.28280
|
| [16] |
Naylor PH, Mutchnick MG (2018) Immunotherapy for hepatitis B in the direct acting antiviral era: reevaluating the thymosin α1 efficacy trials in the light of a combination therapy approach. J Viral Hepat 25: 4–9. doi: 10.1111/jvh.12807
|
| [17] |
Xao G, Yang J, Yan L (2015) Comparison of diagnostic accuracy of aspartate aminotransferase to platelet ration index and fibrosis-4 index for detecting liver fibrosis in adult patients with chronic hepatitis B virus infection: A systemic review and meta-analysis. Hepatology 61: 292–302. doi: 10.1002/hep.27382
|
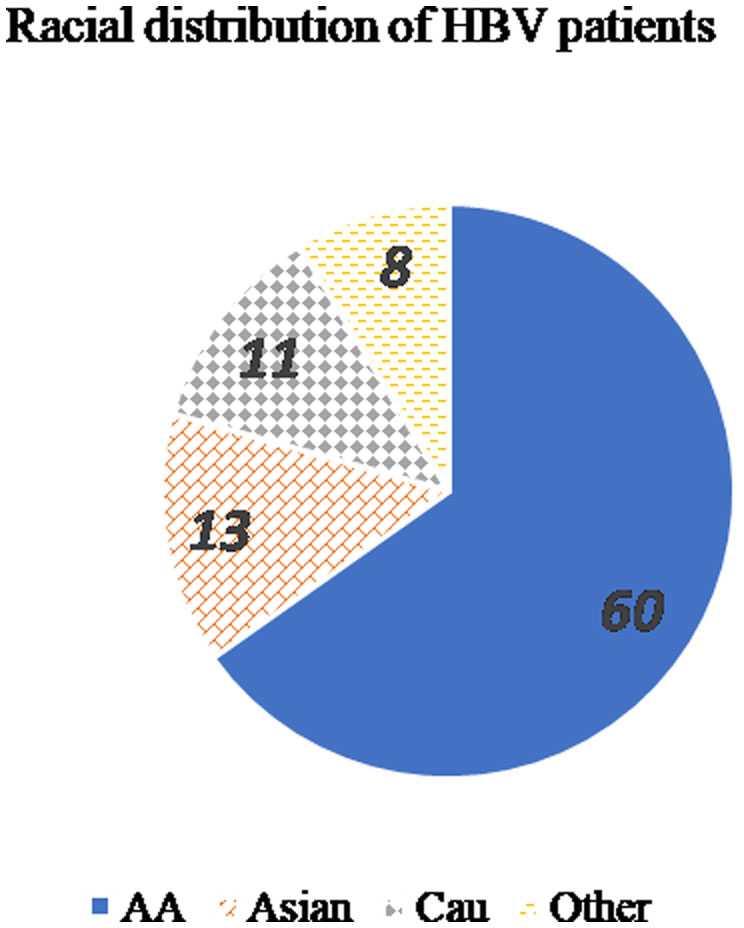
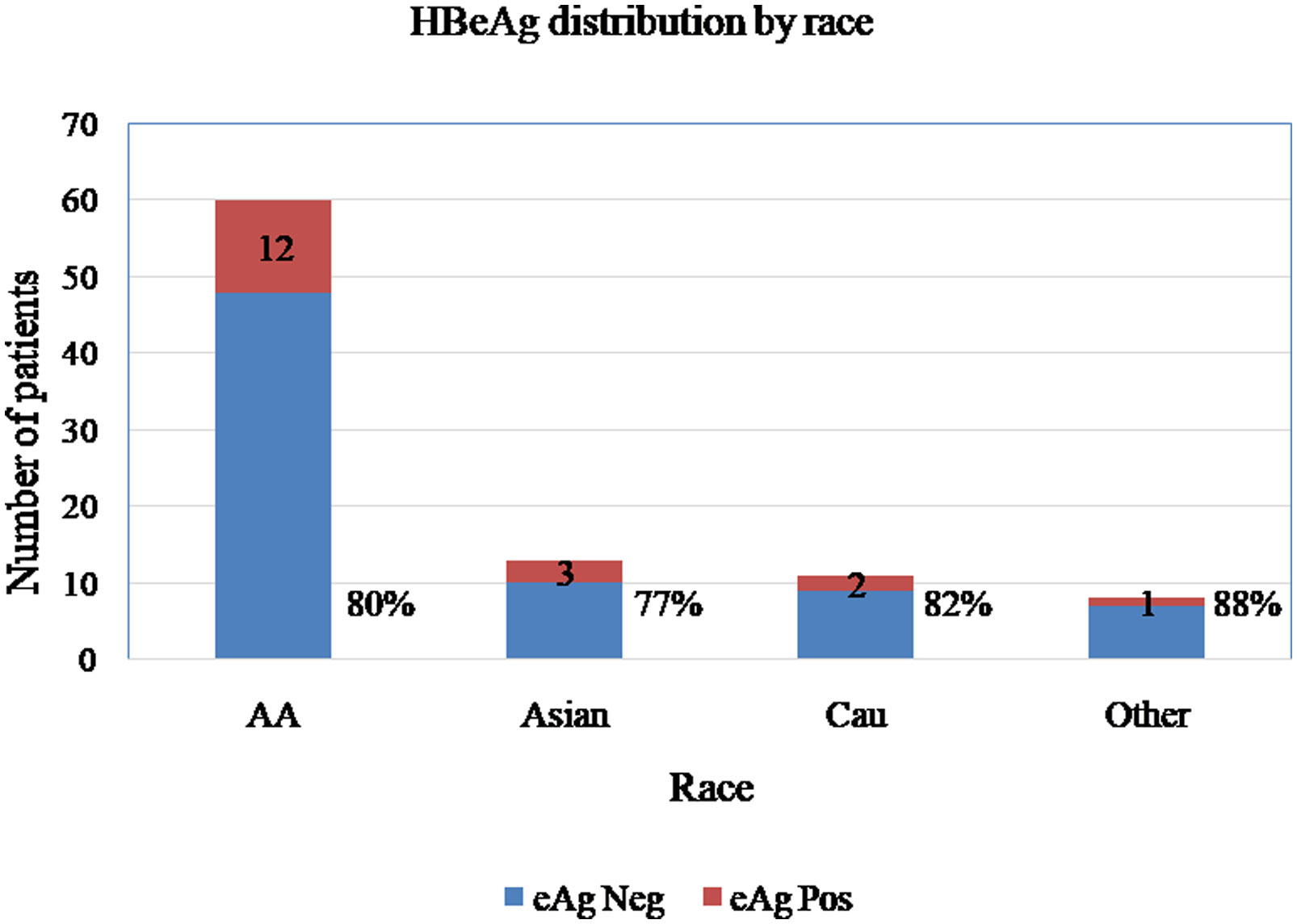
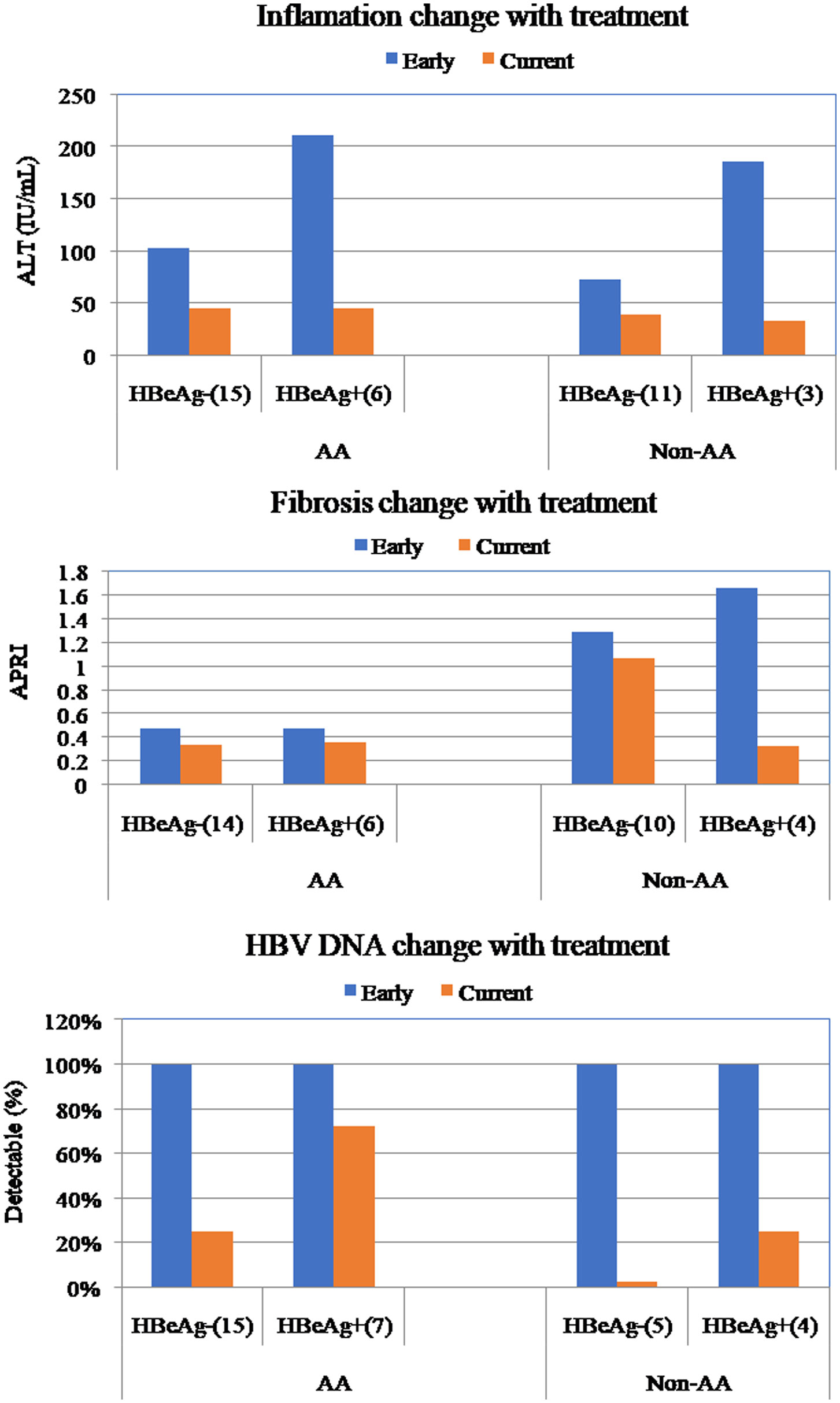
Figures(3) / Tables(2)

Eugene Verkhovsky, Paul Naylor, Sindhuri Benjaram, Shanker Das Kundumadam, Murray Ehrinpreis, Milton Mutchnick. Racial disparity in chronic hepatitis B infection in a predominately African American urban clinic population[J]. AIMS Medical Science, 2018, 5(4): 378-385. doi: 10.3934/medsci.2018.4.378







 DownLoad:
DownLoad: