Citation: Maryam Tanhapour, Asad Vaisi-Raygani, Mozafar Khazaei, Zohreh Rahimi, Tayebeh Pourmotabbed. Cytotoxic T-lymphocyte Associated Antigen-4 (CTLA-4) Polymorphism, Cancer, and Autoimmune Diseases[J]. AIMS Medical Science, 2017, 4(4): 395-412. doi: 10.3934/medsci.2017.4.395

| [1] |
Caspi RR (2008) Immunotherapy of autoimmunity and cancer: The penalty for success. Nat Rev Immunol 8: 970-976. doi: 10.1038/nri2438

|
| [2] | Leach DR, Krummel MF, Allison JP (1996) Enhancement of antitumor immunity by ctla-4 blockade. Science 271: 1734. |
| [3] | Janeway Jr CA, Travers P, Walport M, et al. (2001) The major histocompatibility complex and its functions. Immunobiology. |
| [4] | Goldrath AW, Bevan MJ (1999) Selecting and maintaining a diverse t-cell repertoire. Nature 402: 255-262. |
| [5] |
Chambers CA, Kuhns MS, Egen JG, et al. (2001) Ctla-4-mediated inhibition in regulation of t cell responses: Mechanisms and manipulation in tumor immunotherapy. Annu Rev Immunol 19: 565-594. doi: 10.1146/annurev.immunol.19.1.565
|
| [6] | Rudd CE, Taylor A, Schneider H (2009) Cd28 and ctla‐4 coreceptor expression and signal transduction. Immunol Rev 229: 12-26. |
| [7] | Rutkowski R, Moniuszko T, Stasiak-Barmuta A, et al. (2003) Cd80 and cd86 expression on lps-stimulated monocytes and the effect of cd80 and cd86 blockade on il-4 and ifn-gamma production in nanotopic bronchial asthma. Arch Immunol Ther Exp (Warsz) 51: 421-428. |
| [8] |
Linsley PS, Clark EA, Ledbetter JA (1990) T-cell antigen cd28 mediates adhesion with b cells by interacting with activation antigen b7/bb-1. Proc Natl Acad Sci U S A 87: 5031-5035. doi: 10.1073/pnas.87.13.5031
|
| [9] |
Greenwald RJ, Freeman GJ, Sharpe AH (2005) The b7 family revisited. Annu Rev Immunol 23: 515-548. doi: 10.1146/annurev.immunol.23.021704.115611
|
| [10] | Collins AV, Brodie DW, Gilbert RJ, et al. (2002) The interaction properties of costimulatory molecules revisited. Immunity 17: 201-210. |
| [11] | Sansom D (2000) Cd28, ctla‐4 and their ligands: Who does what and to whom? Immunology 101: 169-177. |
| [12] |
Wang XB, Zheng CY, Giscombe R, et al. (2001) Regulation of surface and intracellular expression of ctla‐4 on human peripheral t cells. Scand J Immunol 54: 453-458. doi: 10.1046/j.1365-3083.2001.00985.x
|
| [13] |
Linsley PS, Greene J, Tan P, et al. (1992) Coexpression and functional cooperation of ctla-4 and cd28 on activated t lymphocytes. J Exp Med 176: 1595-1604. doi: 10.1084/jem.176.6.1595
|
| [14] | Buc M (1996) Immunopathogenic mechanisms in autoimmune processes: Autoantigens. Bratisl Lek Listy 97: 187-195. |
| [15] |
Ribas A, Camacho LH, Lopez-Berestein G, et al. (2005) Antitumor activity in melanoma and anti-self responses in a phase i trial with the anti-cytotoxic t lymphocyte–associated antigen 4 monoclonal antibody cp-675,206. J Clin Oncol 23: 8968-8977. doi: 10.1200/JCO.2005.01.109
|
| [16] |
Maker AV, Phan GQ, Attia P, et al. (2005) Tumor regression and autoimmunity in patients treated with cytotoxic t lymphocyte–associated antigen 4 blockade and interleukin 2: A phase i/ii study. Ann Surg Oncol 12: 1005-1016. doi: 10.1245/ASO.2005.03.536
|
| [17] |
Buchbinder EI, Desai A (2016) Ctla-4 and pd-1 pathways: Similarities, differences, and implications of their inhibition. Am J Clin Oncol 39: 98. doi: 10.1097/COC.0000000000000239
|
| [18] |
Quezada SA, Peggs KS, Curran MA, et al. (2006) Ctla4 blockade and gm-csf combination immunotherapy alters the intratumor balance of effector and regulatory t cells. J Clin Invest 116: 1935-1945. doi: 10.1172/JCI27745
|
| [19] | Phan GQ, Yang JC, Sherry RM, et al. (2003) Cancer regression and autoimmunity induced by cytotoxic t lymphocyte-associated antigen 4 blockade in patients with metastatic melanoma. Proc Natl Acad Sci U S A 100: 8372-8377. |
| [20] |
Hodi FS, Mihm MC, Soiffer RJ, et al. (2003) Biologic activity of cytotoxic t lymphocyte-associated antigen 4 antibody blockade in previously vaccinated metastatic melanoma and ovarian carcinoma patients. Proc Natl Acad Sci U S A 100: 4712-4717. doi: 10.1073/pnas.0830997100
|
| [21] |
Ribas A, Glaspy JA, Lee Y, et al. (2004) Role of dendritic cell phenotype, determinant spreading, and negative costimulatory blockade in dendritic cell-based melanoma immunotherapy. J Immunother 27: 354-367. doi: 10.1097/00002371-200409000-00004
|
| [22] | Wolchok JD, Saenger Y (2008) The mechanism of anti-ctla-4 activity and the negative regulation of t-cell activation. Oncologist 13: 2-9. |
| [23] |
Cooper GS, Miller FW, Pandey JP (1999) The role of genetic factors in autoimmune disease: Implications for environmental research. Environ Health Perspect 107: 693. doi: 10.1289/ehp.99107s5693
|
| [24] | Remmersl EF, Longmanl RE, Dul Y, et al. (1996) A genome scan localizes five non-mhc loci controlling. Nat Genet 14. |
| [25] |
Gupta B, Hawkins RD (2015) Epigenomics of autoimmune diseases. Immunol Cell Biol 93: 271-276. doi: 10.1038/icb.2015.18
|
| [26] | Maurano MT, Humbert R, Rynes E, et al. (2012) Systematic localization of common disease-associated variation in regulatory DNA. Science 337: 1190-1195. |
| [27] |
Kamel AM, Mira MF, Mossallam GI, et al. (2014) Lack of association of ctla-4 +49 a/g polymorphism with predisposition to type 1 diabetes in a cohort of egyptian families. Egypt J Med Hum Genet 15: 25-30. doi: 10.1016/j.ejmhg.2013.09.002
|
| [28] |
Steiner K, Moosig F, Csernok E, et al. (2001) Increased expression of ctla‐4 (cd152) by t and b lymphocytes in wegener's granulomatosis. Clin Exp Immunol 126: 143-150. doi: 10.1046/j.1365-2249.2001.01575.x
|
| [29] |
Teft WA, Kirchhof MG, Madrenas J (2006) A molecular perspective of ctla-4 function. Annu Rev Immunol 24: 65-97. doi: 10.1146/annurev.immunol.24.021605.090535
|
| [30] | Prans E (2010) Allelic variants of ctla-4 gene as important markers of immune regulation in type 1 diabetes. |
| [31] | Pawlak E, Kochanowska IE, Frydecka I, et al. (2005) The soluble ctla-4 receptor: A new marker in autoimmune diseases. Arch Immunol Ther Exp (Warsz) 53: 336. |
| [32] |
Ueda H, Howson JM, Esposito L, et al. (2003) Association of the t-cell regulatory gene ctla4 with susceptibility to autoimmune disease. Nature 423: 506-511. doi: 10.1038/nature01621
|
| [33] |
Chistiakov D, Turakulov R (2003) Ctla-4 and its role in autoimmune thyroid disease. J Mol Endocrinol 31: 21-36. doi: 10.1677/jme.0.0310021
|
| [34] |
Magistrelli G, Jeannin P, Herbault N, et al. (1999) A soluble form of ctla‐4 generated by alternative splicing is expressed by nonstimulated human t cells. Eur J Immunol 29: 3596-3602. doi: 10.1002/(SICI)1521-4141(199911)29:11<3596::AID-IMMU3596>3.0.CO;2-Y
|
| [35] | Jakubczik F, Jones K, Nichols J, et al. (2016) A snp in the immunoregulatory molecule ctla-4 controls mrna splicing in vivo but does not alter diabetes susceptibility in the nod mouse. Diabetes 65: 120-128. |
| [36] | Ghaderi A (2011) Ctla4 gene variants in autoimmunity and cancer: A comparative review. Iran J Immunol 8: 127. |
| [37] |
Valk E, Rudd CE, Schneider H (2008) Ctla-4 trafficking and surface expression. Trends Immunol 29: 272-279. doi: 10.1016/j.it.2008.02.011
|
| [38] |
Gerold KD, Zheng P, Rainbow DB, et al. (2011) The soluble ctla-4 splice variant protects from type 1 diabetes and potentiates regulatory t-cell function. Diabetes 60: 1955-1963. doi: 10.2337/db11-0130
|
| [39] |
Rudd CE (2008) The reverse stop-signal model for ctla4 function. Nat Rev Immunol 8: 153-160. doi: 10.1038/nri2253
|
| [40] |
Walker LS, Sansom DM (2011) The emerging role of ctla4 as a cell-extrinsic regulator of t cell responses. Nat Rev Immunol 11: 852-863. doi: 10.1038/nri3108
|
| [41] |
Parry RV, Chemnitz JM, Frauwirth KA, et al. (2005) Ctla-4 and pd-1 receptors inhibit t-cell activation by distinct mechanisms. Mol Cell Biol 25: 9543-9553. doi: 10.1128/MCB.25.21.9543-9553.2005
|
| [42] |
Chikuma S, Imboden JB, Bluestone JA (2003) Negative regulation of t cell receptor–lipid raft interaction by cytotoxic t lymphocyte–associated antigen 4. J Exp Med 197: 129-135. doi: 10.1084/jem.20021646
|
| [43] |
Choi JM, Ahn MH, Chae WJ, et al. (2006) Intranasal delivery of the cytoplasmic domain of ctla-4 using a novel protein transduction domain prevents allergic inflammation. Nat Med 12: 574-579. doi: 10.1038/nm1385
|
| [44] | Thompson CB, Allison JP (1997) The emerging role of ctla-4 as an immune attenuator. Immunity 7: 445-450. |
| [45] |
Schneider H, Downey J, Smith A, et al. (2006) Reversal of the tcr stop signal by ctla-4. Science 313: 1972-1975. doi: 10.1126/science.1131078
|
| [46] |
Uyttenhove C, Pilotte L, Théate I, et al. (2003) Evidence for a tumoral immune resistance mechanism based on tryptophan degradation by indoleamine 2, 3-dioxygenase. Nat med 9: 1269-1274. doi: 10.1038/nm934
|
| [47] |
Hwang SL, Chung NP-y, Chan JK-y, et al. (2005) Indoleamine 2, 3-dioxygenase (ido) is essential for dendritic cell activation and chemotactic responsiveness to chemokines. Cell res 15: 167-175. doi: 10.1038/sj.cr.7290282
|
| [48] |
Munn DH, Mellor AL (2007) Indoleamine 2, 3-dioxygenase and tumor-induced tolerance. J Clin Invest 117: 1147-1154. doi: 10.1172/JCI31178
|
| [49] |
Vignali DA, Collison LW, Workman CJ (2008) How regulatory t cells work. Nat Rev Immunol 8: 523-532. doi: 10.1038/nri2343
|
| [50] | Gorelik L, Flavell RA (2000) Abrogation of tgfβ signaling in t cells leads to spontaneous t cell differentiation and autoimmune disease. Immunity 12: 171-181. |
| [51] |
Qureshi OS, Zheng Y, Nakamura K, et al. (2011) Trans-endocytosis of cd80 and cd86: A molecular basis for the cell-extrinsic function of ctla-4. Science 332: 600-603. doi: 10.1126/science.1202947
|
| [52] |
Carreno BM, Bennett F, Chau TA, et al. (2000) Ctla-4 (cd152) can inhibit t cell activation by two different mechanisms depending on its level of cell surface expression. J Immunol 165: 1352-1356. doi: 10.4049/jimmunol.165.3.1352
|
| [53] |
Sharpe AH, Freeman GJ (2002) The b7–cd28 superfamily. Nat Rev Immunol 2: 116-126. doi: 10.1038/nri727
|
| [54] |
Krummey SM, Ford ML (2014) Braking bad: Novel mechanisms of ctla‐4 inhibition of t cell responses. Am J Transplant 14: 2685-2690. doi: 10.1111/ajt.12938
|
| [55] | Grosso JF, Jure-Kunkel MN (2013) Ctla-4 blockade in tumor models: An overview of preclinical and translational research. Cancer Immun 13: 5. |
| [56] |
Schreiber RD, Old LJ, Smyth MJ (2011) Cancer immunoediting: Integrating immunity's roles in cancer suppression and promotion. Science 331: 1565-1570. doi: 10.1126/science.1203486
|
| [57] |
Palacios R, Comas D, Elorza J, et al. (2008) Genomic regulation of ctla4 and multiple sclerosis. J Neuroimmunol 203: 108-115. doi: 10.1016/j.jneuroim.2008.06.021
|
| [58] | Ramirez SA, Lao O, Soldevila M, et al. (2005) Haplotype tagging efficiency in worldwide populations in ctla4 gene. Genes Immun 6: 646-657. |
| [59] |
Zheng J, Yu X, Jiang L, et al. (2010) Association between the cytotoxic t-lymphocyte antigen 4 + 49g> a polymorphism and cancer risk: A meta-analysis. BMC cancer 10: 522. doi: 10.1186/1471-2407-10-522
|
| [60] |
Pérez-García A, De la Cámara R, Román-Gómez J, et al. (2007) Ctla-4 polymorphisms and clinical outcome after allogeneic stem cell transplantation from hla-identical sibling donors. Blood 110: 461-467. doi: 10.1182/blood-2007-01-069781
|
| [61] | Tanhapour M, Vaisi-Raygani A, Bahrehmand F, et al. (2016) Association between the cytotoxic t-lymphocyte antigen-4 mutations and the susceptibility to systemic lupus erythematosus; contribution markers of inflammation and oxidative stress. Cell Mol Biol (Noisy-le-grand) 62: 56. |
| [62] | Ling V, Wu PW, Finnerty HF, et al. (1999) Complete sequence determination of the mouse and human ctla4 gene loci: Cross-species DNA sequence similarity beyond exon borders. Genomics 60: 341-355. |
| [63] | Yanagawa T, Hidaka Y, Guimaraes V, et al. (1995) Ctla-4 gene polymorphism associated with graves' disease in a caucasian population. J Clin Endocrinol Metab 80: 41-45. |
| [64] |
Vaidya B, Pearce S (2004) The emerging role of the ctla-4 gene in autoimmune endocrinopathies. Eur J Endocrinol 150: 619-626. doi: 10.1530/eje.0.1500619
|
| [65] |
Ting WH, Chien MN, Lo FS, et al. (2016) Association of cytotoxic t-lymphocyte-associated protein 4 (ctla4) gene polymorphisms with autoimmune thyroid disease in children and adults: Case-control study. PloS one 11: e0154394. doi: 10.1371/journal.pone.0154394
|
| [66] | Paula AV, Lourdes C, RicardoV GM, et al. (2011) Association of ctla4 gene polymorphism with ophthalmopathy of graves' disease in a spanish population. Int J Endocrinol Metabolism 9: 397-402. |
| [67] | Du P, Ma X, Wang C (2014) Associations of ctla4 gene polymorphisms with graves' ophthalmopathy: A meta-analysis. Int J Genomics. |
| [68] |
Khalilzadeh O, Amiri HM, Tahvildari M, et al. (2009) Pretibial myxedema is associated with polymorphism in exon 1 of ctla-4 gene in patients with graves' ophthalmopathy. Arch Dermatol Res 301: 719-723. doi: 10.1007/s00403-008-0919-1
|
| [69] |
Han S, Zhang S, Zhang W, et al. (2006) Ctla4 polymorphisms and ophthalmopathy in graves' disease patients: Association study and meta-analysis. Hum Immunol 67: 618-626. doi: 10.1016/j.humimm.2006.05.003
|
| [70] |
Si X, Zhang X, Tang W, et al. (2012) Association between the ctla-4 +49a/g polymorphism and graves' disease: A meta-analysis. Exp Ther Med 4: 538-544. doi: 10.3892/etm.2012.618
|
| [71] |
Devaraju P, Gulati R, Singh B, et al. (2014) The ctla4 +49 a/g (rs231775) polymorphism influences susceptibility to sle in south indian tamils. Tissue Antigens 83: 418-421. doi: 10.1111/tan.12363
|
| [72] |
Liu J, Zhang H-X (2013) Ctla-4 polymorphisms and systemic lupus erythematosus: A comprehensive meta-analysis. Genet Test Mol Biomarkers 17: 226-231. doi: 10.1089/gtmb.2012.0302
|
| [73] | Kimkong I, Nakkuntod J, Sae-Ngow S, et al. (2011) Association between ctla-4 polymorphisms and the susceptibility to systemic lupus erythematosus and graves' disease in thai population. Asian Pac J Allergy Immunol 29: 229. |
| [74] |
Chua KH, Puah SM, Chew CH, T et al. (2010) Study of the ctla-4 gene polymorphisms in systemic lupus erythematosus (sle) samples from malaysia. Ann Hum Biol 37: 275-281. doi: 10.3109/03014460903325185
|
| [75] | Ahmed S, Ihara K, Kanemitsu S, et al. (2001) Association of ctla‐4 but not cd28 gene polymorphisms with systemic lupus erythematosus in the japanese population. Rheumatology 40: 662-667. |
| [76] |
Katkam SK, Kumaraswami K, Rupasree Y, T et al. (2016). Association of ctla4 exon-1 polymorphism with the tumor necrosis factor-α in the risk of systemic lupus erythematosus among south indians. Hum Immunol 77: 158-164. doi: 10.1016/j.humimm.2015.11.002
|
| [77] |
Barreto M, Santos E, Ferreira R, et al. (2004) Evidence for ctla4 as a susceptibility gene for systemic lupus erythematosus. Eur J Hum Genet 12: 620-626. doi: 10.1038/sj.ejhg.5201214
|
| [78] |
Zhai JX, Zou LW, Zhang ZX, et al. (2013) Ctla-4 polymorphisms and systemic lupus erythematosus (sle): A meta-analysis. Mol Biol Rep 40: 5213-5223. doi: 10.1007/s11033-012-2125-7
|
| [79] |
Narooie NM, Taji O, Tamandani DMK, et al. (2017) Association of CTLA-4 gene polymorphisms−318c/t and +49a/g and hashimoto's thyroidits in zahedan, iran. Biomed Rep 6: 108-112. doi: 10.3892/br.2016.813
|
| [80] |
Liu J, Zhang HX (2014) Ctla-4 gene and the susceptibility of multiple sclerosis: An updated meta-analysis study including 12,916 cases and 15,455 controls. J Neurogenet 28: 153-163. doi: 10.3109/01677063.2014.880703
|
| [81] |
Farra C, Awwad J, Fadlallah A, et al. (2012) Genetics of autoimmune thyroid disease in the lebanese population. J Community Genet 3: 259-264. doi: 10.1007/s12687-012-0085-1
|
| [82] | Bicek A, Zaletel K, Gaberscek S, et al. (2009) 49a/g and ct60 polymorphisms of the cytotoxic t-lymphocyte-associated antigen 4 gene associated with autoimmune thyroid disease. Hum Immunol 70: 820-824. |
| [83] | Sameem M, Rani A, Bashir R, et al. (2015) Ctla-4 +49 polymorphism and susceptibility to rheumatoid arthritis in pakistani population. Pakistan J Zool 47: 1731-1737. |
| [84] |
Li G, Shi F, Liu J, et al. (2014) The effect of ctla-4 a49g polymorphism on rheumatoid arthritis risk: A meta-analysis. Diagn Pathol 9: 157. doi: 10.1186/s13000-014-0157-0
|
| [85] | Vaidya B, Pearce S, Charlton S, et al. (2002) An association between the ctla4 exon 1 polymorphism and early rheumatoid arthritis with autoimmune endocrinopathies. Rheumatology 41: 180-183. |
| [86] | Dai Z, Tian T, Wang M, et al. (2017) Ctla-4 polymorphisms associate with breast cancer susceptibility in asians: A meta-analysis. Peer J 5: e2815. |
| [87] | Liu P, Xu L, Sun Y, et al. (2014) The association between cytotoxic t lymphocyte-associated antigen-4 and cervical cancer. Tumor Biol 35: 2893-2903. |
| [88] | Han W-JW (2016) Association of cytotoxic t-lymphocyte antigen-4 polymorphisms with malignant bone tumors risk: A meta-analysis. Asian Pac J Cancer Prev 17: 3785-3791. |
| [89] | Sáenz LP, Vázquez AF, Romero JM, et al. (2009) Polymorphisms in inflammatory response genes in metastatic renal cancer. Actas Urol Esp 33: 474-481. |
| [90] |
Jaiswal PK, Singh V, Mittal RD (2014) Cytotoxic t lymphocyte antigen 4 (ctla4) gene polymorphism with bladder cancer risk in north indian population. Mol Biol Rep 41: 799-807. doi: 10.1007/s11033-013-2919-2
|
| [91] |
Hu L, Liu J, Chen X, et al. (2010) Ctla-4 gene polymorphism +49 a/g contributes to genetic susceptibility to two infection-related cancers-hepatocellular carcinoma and cervical cancer. Hum Immunol 71: 888-891. doi: 10.1016/j.humimm.2010.05.023
|
| [92] |
Cheng TY, Lin JT, Chen LT, et al. (2006) Association of t-cell regulatory gene polymorphisms with susceptibility to gastric mucosa-associated lymphoid tissue lymphoma. J Clin Oncol 24: 3483-3489. doi: 10.1200/JCO.2005.05.5434
|
| [93] |
Gao X, Zhang S, Qiao X, et al. (2014) Association of cytotoxic t lymphocyte antigen-4 +49a/g polymorphism and cancer risk: An updated meta-analysis. Cancer Biomark 14: 287-294. doi: 10.3233/CBM-140403
|
| [94] | Minhas S, Bhalla S, Shokeen Y, et al. (2014) Lack of any association of the ctla-4 +49 g/a polymorphism with breast cancer risk in a north indian population. Asian Pac J Cancer Prev 15: 2035-2038. |
| [95] | Qiu H, Tang W, Yin P, et al. (2013) Cytotoxic t-lymphocyte-associated antigen‑4 polymorphisms and susceptibility to cervical cancer: A meta-analysis. Mol Med Rep 8: 1785-1794. |
| [96] |
Khaghanzadeh N, Erfani N, Ghayum MA, et al. (2010) Ctla4 gene variations and haplotypes in patients with lung cancer. Cancer Genet Cytogenet 196: 171-174. doi: 10.1016/j.cancergencyto.2009.09.001
|
| [97] |
Su T-H, Chang T-Y, Lee Y-J, et al. (2007) Ctla-4 gene and susceptibility to human papillomavirus-16-associated cervical squamous cell carcinoma in taiwanese women. Carcinogenesis 28: 1237-1240. doi: 10.1093/carcin/bgm043
|
| [98] |
Bharti V, Mohanti BK, Das SN (2013) Functional genetic variants of ctla-4 and risk of tobacco-related oral carcinoma in high-risk north indian population. Hum Immunol 74: 348-352. doi: 10.1016/j.humimm.2012.12.008
|
| [99] | Touma Z, Hamdan A, Shamseddeen W, et al. (2008) CTLA-4 gene variants are not associated with Behçet's disease or its clinical manifestations. Clin Exp Rheumatol 26: S132. |
| [100] |
Wang L, Li D, Fu Z, et al. (2007) Association of ctla-4 gene polymorphisms with sporadic breast cancer in chinese han population. BMC cancer 7: 173. doi: 10.1186/1471-2407-7-173
|
| [101] | Perez-Garcia A, Brunet S, Berlanga J, et al. (2009) Ctla-4 genotype and relapse incidence in patients with acute myeloid leukemia in first complete remission after induction chemotherapy. Leukemia 23: 486-491. |
| [102] | Erfani N, Haghshenas MR, Hoseini MA, et al. (2012) Strong association of ctla-4 variation (ct60a/g) and ctla-4 haplotypes with predisposition of iranians to head and neck cancer. Iran J Immunol 9: 188. |
| [103] |
Torres B, Aguilar F, Franco E, et al. (2004) Association of the ct60 marker of the ctla4 gene with systemic lupus erythematosus. Arthritis Rheum 50: 2211-2215. doi: 10.1002/art.20347
|
| [104] | Lu L, Wang W, Feng R, et al. (2016) Association between cytotoxic t lymphocyte antigen-4 gene polymorphisms and gastric cancer risk: A meta-analysis of case-control studies. Int J Clin Exp Med 9: 10639-10650. |
| [105] |
Chong KK, Chiang SW, Wong GW, et al. (2008) Association of ctla-4 and il-13 gene polymorphisms with graves' disease and ophthalmopathy in chinese children. Invest Ophthalmol Vis Sci 49: 2409-2415. doi: 10.1167/iovs.07-1433
|
| [106] | Tomoyose T, Komiya I, Takara M, et al. (2002) Cytotoxic t-lymphocyte antigen-4 gene polymorphisms and human t-cell lymphotrophic virus-1 infection: Their associations with hashimoto's thyroiditis in japanese patients. Thyroid 12: 673-677. |
| [107] |
Hudson LL, Rocca K, Song YW, et al. (2002) Ctla-4 gene polymorphisms in systemic lupus erythematosus: A highly significant association with a determinant in the promoter region. Hum Genet 111: 452-455. doi: 10.1007/s00439-002-0807-2
|
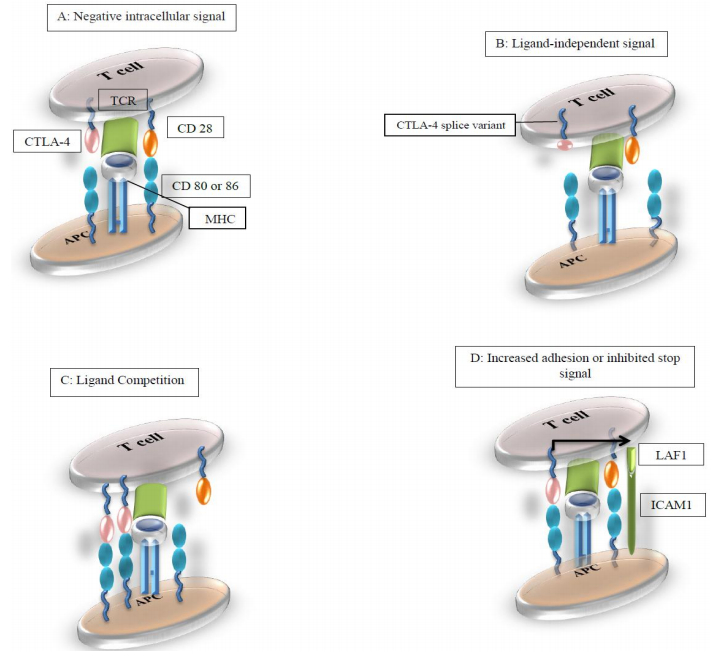
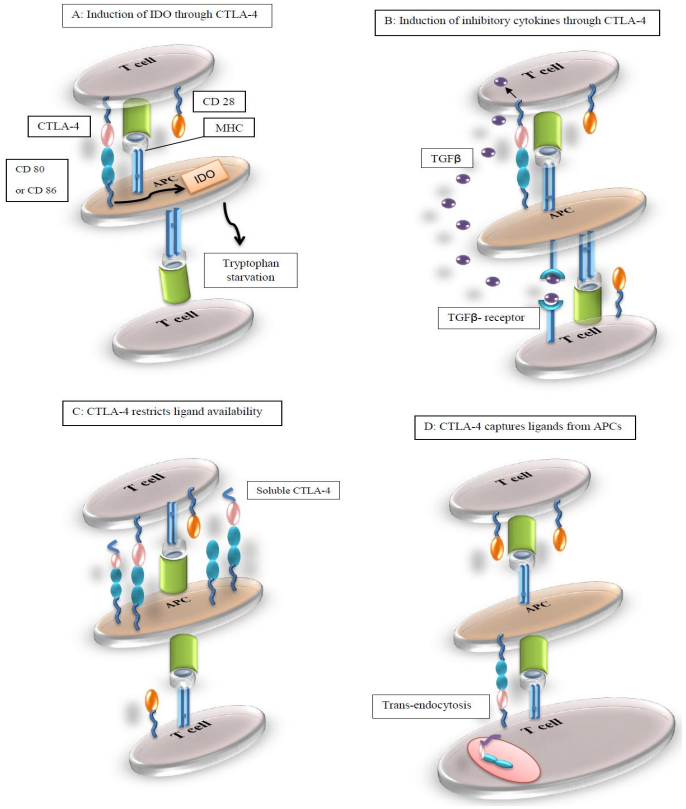
Figures(2) / Tables(2)

Maryam Tanhapour, Asad Vaisi-Raygani, Mozafar Khazaei, Zohreh Rahimi, Tayebeh Pourmotabbed. Cytotoxic T-lymphocyte Associated Antigen-4 (CTLA-4) Polymorphism, Cancer, and Autoimmune Diseases[J]. AIMS Medical Science, 2017, 4(4): 395-412. doi: 10.3934/medsci.2017.4.395







 DownLoad:
DownLoad: