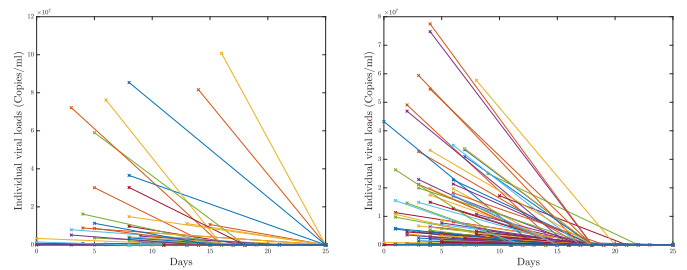
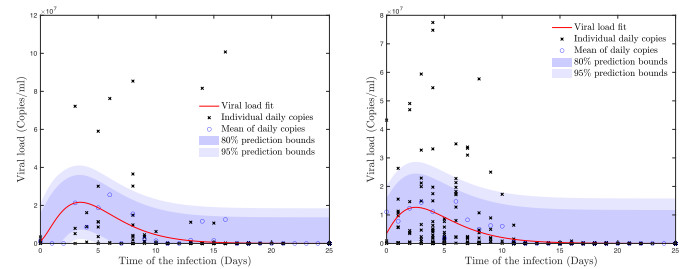
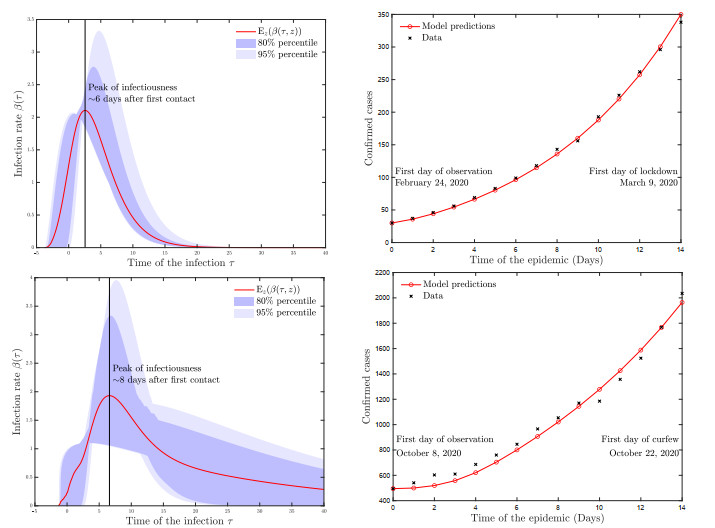
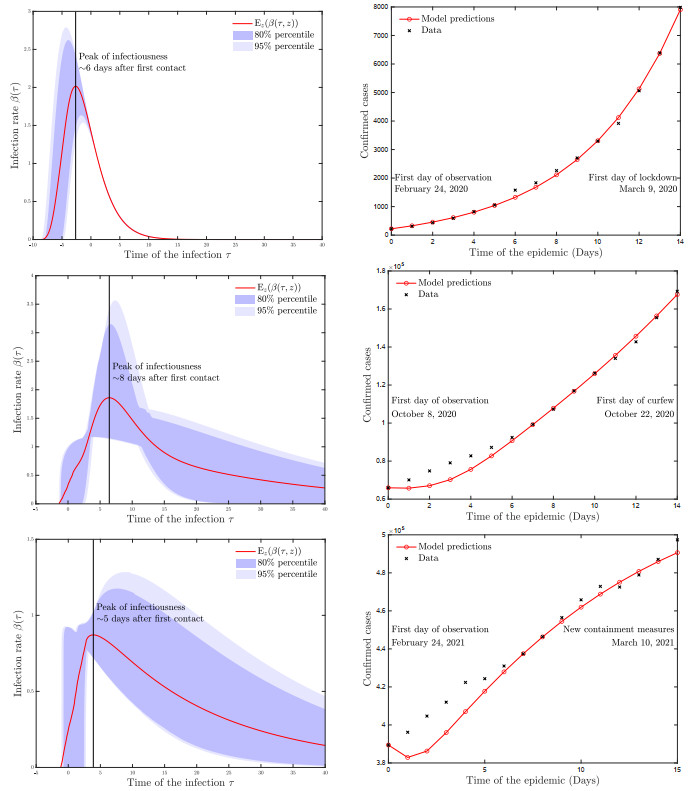
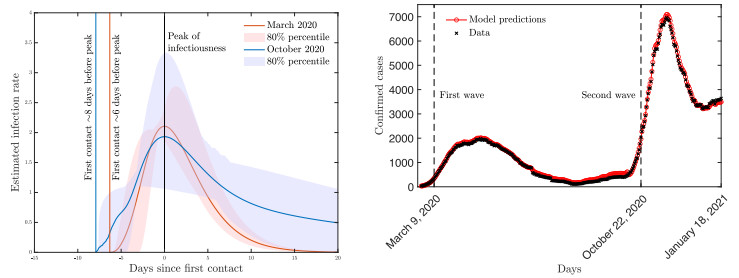
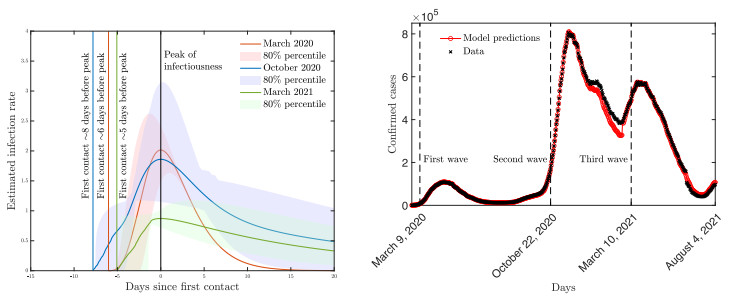
A pipeline to evaluate the evolution of viral dynamics based on a new model-driven approach has been developed in the present study. The proposed methods exploit real data and the multiscale structure of the infection dynamics to provide robust predictions of the epidemic dynamics. We focused on viral load kinetics whose dynamical features are typically available in the symptomatic stage of the infection. Hence, the epidemiological evolution was obtained by relying on a compartmental approach characterized by a varying infection rate to estimate early-stage viral load dynamics, of which few data are available. We tested the proposed approach with real data of SARS-CoV-2 viral load kinetics collected from patients living in an Italian province. The considered database refers to early-phase infections, whose viral load kinetics have not been affected by the mass vaccination policies in Italy.
Citation: Andrea Bondesan, Antonio Piralla, Elena Ballante, Antonino Maria Guglielmo Pitrolo, Silvia Figini, Fausto Baldanti, Mattia Zanella. Predictability of viral load dynamics in the early phases of SARS-CoV-2 through a model-based approach[J]. Mathematical Biosciences and Engineering, 2025, 22(4): 725-743. doi: 10.3934/mbe.2025027
A pipeline to evaluate the evolution of viral dynamics based on a new model-driven approach has been developed in the present study. The proposed methods exploit real data and the multiscale structure of the infection dynamics to provide robust predictions of the epidemic dynamics. We focused on viral load kinetics whose dynamical features are typically available in the symptomatic stage of the infection. Hence, the epidemiological evolution was obtained by relying on a compartmental approach characterized by a varying infection rate to estimate early-stage viral load dynamics, of which few data are available. We tested the proposed approach with real data of SARS-CoV-2 viral load kinetics collected from patients living in an Italian province. The considered database refers to early-phase infections, whose viral load kinetics have not been affected by the mass vaccination policies in Italy.

| [1] |
L. Bolzoni, E. Bonacini, C. Soresina, M. Groppi, Time-optimal control strategies in SIR epidemic models, Math. Biosci., 292 (2017), 86–96. https://doi.org/10.1016/j.mbs.2017.07.011 doi: 10.1016/j.mbs.2017.07.011

|
| [2] | F. Brauer, C. Castillo-Chavez, Z. Feng, Mathematical Models in Epidemiology, in Texts in Applied Mathematics, Springer, New York, 2019. |
| [3] |
T. Britton, F. Ball, P. Trapman, A mathematical model reveals the influence of population heterogeneity on herd immunity to SARS-CoV-2, Science, 369 (2020), 846–849. https://doi.org/10.1126/science.abc6810 doi: 10.1126/science.abc6810
|
| [4] |
B. Buonomo, R. Della Marca, Effects of information-induced behavioural changes during the COVID-19 lockdowns: the case of Italy, R. Soc. Open Sci., 7 (2020), 201635. https://doi.org/10.1098/rsos.201635 doi: 10.1098/rsos.201635
|
| [5] |
J. Chen, H. Lu, G. Melino, S. Boccia, M. Piacentini, W. Ricciardi, et al., COVID-19 infection: the China and Italy perspectives, Cell Death Dis., 11 (2020), 438. https://doi.org/10.1038/s41419-020-2603-0 doi: 10.1038/s41419-020-2603-0
|
| [6] |
M. Gatto, E. Bertuzzo, L. Mari, S. Miccoli, L. Carraro, R. Casagrandi, et al., Spread and dynamics of the COVID-19 epidemic in Italy: effect of emergency containment measures, PNAS, 117 (2020), 10484–10491. https://doi.org/10.1073/pnas.2004978117 doi: 10.1073/pnas.2004978117
|
| [7] |
G. Giordano, F. Blanchini, R. Bruno, P. Colaneri, A. Di Filippo, A. Di Matteo, et al., Modelling the COVID-19 epidemic and implementation of population-wide interventions in Italy, Nat. Med., 26 (2020), 855–860. https://doi.org/10.1038/s41591-020-0883-7 doi: 10.1038/s41591-020-0883-7
|
| [8] | M. J. Keeling, L. Dyson, M. J. Tildesley, E. M. Hill, S. Moore, Comparison of the 2021 COVID-19 roadmap projections against public health data in England, Nat. Commun., 13 (2022). https://doi.org/10.1038/s41467-022-31991-0 |
| [9] |
O. Puhach, B. Meyer, I. Eckerle, SARS-CoV-2 viral load and shedding kinetics, Nat. Rev. Microbiol., 21 (2023), 147–161. https://doi.org/10.1038/s41579-022-00822-w doi: 10.1038/s41579-022-00822-w
|
| [10] |
M. Zanella, C. Bardelli, G. Dimarco, S. Deandrea, P. Perotti, M. Azzi, et al., A data-driven epidemic model with social structure for understanding the COVID-19 infection on a heavily affected Italian Province, Math. Models Methods Appl. Sci., 31 (2023), 2533–2570. https://doi.org/10.1142/S021820252150055X doi: 10.1142/S021820252150055X
|
| [11] |
T. C. Jones, G. Biele, B. Mühlemann, T. Veith, J. Schneider, J. Beheim-Schwarzbach, et al., Estimating infectiousness throughout SARS-CoV-2 infection course, Science, 373 (2021), eabi5273. https://doi.org/10.1126/science.abi5273 doi: 10.1126/science.abi5273
|
| [12] |
H. Lu, F. Giannino, D. M. Tartakovsky, Parsimonious models of in-host viral dynamics and immune response, Appl. Math. Lett., 145 (2023), 108781. https://doi.org/10.1016/j.aml.2023.108781 doi: 10.1016/j.aml.2023.108781
|
| [13] |
G. Bertaglia, L. Pareschi, G. Toscani, Modelling contagious viral dynamics: a kinetic approach based on mutual utility, Math. Biosci. Eng., 21 (2024), 4241–4268. https://doi.org/10.3934/mbe.2024187 doi: 10.3934/mbe.2024187
|
| [14] |
J. K. Frediani, R. Parsons, K. B. McLendon, A. L. Westbrook, W. Lam, G. Martin, et al., The new normal: delayed peak SARS-CoV-2 viral loads relative to symptom onset and implications for COVID-19 testing programs, Clin. Infect. Dis., 78 (2024), 301–307. https://doi.org/10.1093/cid/ciad582 doi: 10.1093/cid/ciad582
|
| [15] |
O. Puhach, K. Adea, N. Hulo, P. Sattonnet, C. Genecand, A. Iten, et al., Infectious viral load in unvaccinated and vaccinated individuals infected with ancestral, Delta or Omicron SARS-CoV-2, Nat. Med., 28 (2022), 1491–1500. https://doi.org/10.1038/s41591-022-01816-0 doi: 10.1038/s41591-022-01816-0
|
| [16] |
R. Verity, L. C. Okell, I. Dorigatti, P. Winskill, C. Whittaker, N. Imai, et al., Estimates of the severity of coronavirus disease 2019: a model-based analysis, Lancet Infect. Dis., 20 (2020), 669–677. https://doi.org/10.1016/S1473-3099(20)30243-7 doi: 10.1016/S1473-3099(20)30243-7
|
| [17] |
X. He, E. H. Y. Lau, P. Wu, X. Deng, J. Wang, X. Hao, et al., Temporal dynamics in viral shedding and transmissibility of COVID-19, Nat. Med., 26 (2020), 672–675. https://doi.org/10.1038/s41591-020-0869-5 doi: 10.1038/s41591-020-0869-5
|
| [18] |
E. Kharazmi, M. Cai, X. Zheng, Z. Zhang, G. Lin, G. E. Karniadakis, Identifiability and predictability of integer- and fractional-order epidemiological models using physics-informed neural networks, Nature Comput. Sci., 1 (2021), 744–753. https://doi.org/10.1038/s43588-021-00158-0 doi: 10.1038/s43588-021-00158-0
|
| [19] |
R. Della Marca, N. Loy, A. Tosin, An SIR model with viral load-dependent transmission, J. Math. Biol., 86 (2023), 61. https://doi.org/10.1007/s00285-023-01901-z doi: 10.1007/s00285-023-01901-z
|
| [20] |
N. Loy, A. Tosin, A viral load-based model for epidemic spread on spatial networks, Math. Biosci. Eng., 18 (2021), 5635–5663. https://doi.org/10.3934/mbe.2021285 doi: 10.3934/mbe.2021285
|
| [21] |
G. Bertaglia, A. Bondesan, D. Burini, R. Eftimie, L. Pareschi, G. Toscani, New trends on the systems approach to modeling SARS-CoV-2 pandemics in a globally connected planet, Math. Models Methods Appl. Sci., 34 (2024), 1995–2054. https://doi.org/10.1142/S0218202524500301 doi: 10.1142/S0218202524500301
|
| [22] |
G. Bertaglia, W. Boscheri, G. Dimarco, L. Pareschi, Spatial spread of COVID-19 outbreak in Italy using multiscale kinetic transport equations with uncertainty, Math. Biosci. Eng., 18 (2021), 7028–7059. https://doi.org/10.3934/mbe.2021350 doi: 10.3934/mbe.2021350
|
| [23] |
A. Bondesan, G. Toscani, M. Zanella, Kinetic compartmental models driven by opinion dynamics: Vaccine hesitancy and social influence, Math. Models Methods Appl. Sci., 34 (2024), 1043–1076. https://doi.org/10.1142/S0218202524400062 doi: 10.1142/S0218202524400062
|
| [24] |
G. Dimarco, B. Perthame, G. Toscani, M. Zanella, Kinetic models for epidemic dynamics with social heterogeneity, J. Math. Biol., 83 (2021), 4. https://doi.org/10.1007/s00285-021-01630-1 doi: 10.1007/s00285-021-01630-1
|
| [25] |
J. Franceschi, A. Medaglia, M. Zanella, On the optimal control of kinetic epidemic models with uncertain social features, Optim. Contr. Appl. Meth., 45 (2024), 494–522. https://doi.org/10.1002/oca.3029 doi: 10.1002/oca.3029
|
| [26] |
M. Zanella, Kinetic models for epidemic dynamics in the presence of opinion polarization, Bull. Math. Biol., 85 (2023), 36. https://doi.org/10.1007/s11538-023-01147-2 doi: 10.1007/s11538-023-01147-2
|
| [27] |
J. Demongeot, Q. Griette, Y. Maday, P. Magal, A Kermack–McKendrick model with age of infection starting from a single or multiple cohorts of infected patients, Proc. R. Soc. A, 479 (2023), 20220381. https://doi.org/10.1098/rspa.2022.0381 doi: 10.1098/rspa.2022.0381
|
| [28] |
J. Y. Yang, F. Q. Zhang, X. Y. Wang, SIV epidemic models with age of infection, Int. J. Biomath., 2 (2009), 61–67. https://doi.org/10.1142/S1793524509000492 doi: 10.1142/S1793524509000492
|
| [29] |
A. Piralla, C. Daleno, E. Pariani, P. Conaldi, S. Esposito, A. Zanetti, et al., Virtual quantification of influenza A virus load by real-time RT-PCR, J. Clin. Virol., 56 (2013), 65–68. https://doi.org/10.1016/j.jcv.2012.09.011 doi: 10.1016/j.jcv.2012.09.011
|
| [30] |
A. Piralla, A. Girello, M. Premoli, F. Baldanti, A new real-time reverse transcription-PCR assay for detection of human enterovirus 68 in respiratory samples, J. Clin. Microbiol., 53 (2015), 1725–1726. https://doi.org/10.1128/JCM.03691-14 doi: 10.1128/JCM.03691-14
|
| [31] |
M. Cevik, K. Kuppalli, J. Kindrachuk, M. Peiris, Virology, transmission, and pathogenesis of SARS-CoV-2, BMJ, 371 (2020), 1–6. https://doi.org/10.1136/bmj.m3862 doi: 10.1136/bmj.m3862
|
| [32] | O. Diekmann, J. A. P. Heesterbeek, Mathematical Epidemiology of Infectious Diseases: Model Building, Analysis and Interpretation, Wiley, 2000. |
| [33] |
G. Albi, L. Pareschi, M. Zanella, Control with uncertain data of socially structured compartmental epidemic models, J. Math. Biol., 82 (2021), 63. https://doi.org/10.1007/s00285-021-01617-y doi: 10.1007/s00285-021-01617-y
|
| [34] |
B. Bartélemy, A. Barrat, R. Pastor-Satorras, A. Vespignani, Dynamical patterns of epidemic outbreaks in complex heterogeneous networks, J. Theor. Biol., 235 (2005), 275–288. https://doi.org/10.1016/j.jtbi.2005.01.011 doi: 10.1016/j.jtbi.2005.01.011
|
| [35] |
V. Capasso, G. Serio, A generalization of the Kermack–McKendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43–61. https://doi.org/10.1016/0025-5564(78)90006-8 doi: 10.1016/0025-5564(78)90006-8
|
| [36] |
J. Dolbeault, G. Turinici, Heterogeneous social interactions and the COVID-19 lockdown outcome in a multi-group SEIR model, Math. Model. Net. Pheno., 15 (2020), 1–18. https://doi.org/10.1051/mmnp/2020025 doi: 10.1051/mmnp/2020025
|
| [37] | M. Mozgovoj, M. D. Graham, C. Ferrufino, S. Blanc, A. F. Souto, M. Pilloff, et al., Viral load in symptomatic and asymptomatic patients infected with SARS-CoV-2. What have we learned?, J. Clin. Virol. Plus, 3 (2023), 100166. |
| [38] |
M. A. Capistrán, J. A. Christen, J. X. Velasco-Hernández, Towards uncertainty quantification and inference in the stochastic SIR epidemic model, Math. Biosci., 240 (2012), 250–259. https://doi.org/10.1016/j.mbs.2012.08.005 doi: 10.1016/j.mbs.2012.08.005
|
| [39] | G. Chowell, Fitting dynamic models to epidemic outbreaks with quantified uncertainty: a primer for parameter uncertainty, identifiability, and forecasts, Infect. Dis. Model., 2 (2017), 379–398. |
| [40] |
M. G. Roberts, Epidemic models with uncertainty in the reproduction number, J. Math. Biol., 66 (2013), 1463–1474. https://doi.org/10.1007/s00285-012-0540-y doi: 10.1007/s00285-012-0540-y
|
| [41] | S. Jin, L. Pareschi, Uncertainty Quantification for Hyperbolic and Kinetic Equations, SEMA-SIMAI Springer Series, 2017. |
| [42] | L. Pareschi, An introduction to uncertainty quantification for kinetic equations and related problems, in Trails in Kinetic Theory: Foundational Aspects and Numerical Methods (eds. G. Albi, S. Merino-Aceituno, A. Nota, M. Zanella), SEMA-SIMAI Springer Series, 25 (2021). https://doi.org/10.1007/978-3-030-67104-4 |
| [43] | D. Xiu, Numerical Methods for Stochastic Computations: a Spectral Methods Approach, Princeton University Press, Princeton, 2010. |
| [44] |
N. Guglielmi, E. Iacomini, A. Viguerie, Identification of time delays in COVID-19 data, Epidemiol. Meth., 12 (2023), 20220117. https://doi.org/10.1515/em-2022-0117 doi: 10.1515/em-2022-0117
|
| [45] |
E. Lavezzo, E. Franchin, C. Ciavarella, G. Cuomo-Dannenburg, L. Barzon, C. Del Vecchio, et al., Suppression of a SARS-CoV-2 outbreak in the Italian municipality of Vo', Nature, 584 (2020), 425–429. https://doi.org/10.1038/s41586-020-2488-1 doi: 10.1038/s41586-020-2488-1
|
| [46] |
B. La Scola, M. Le Bideau, J. Andreani, V. T. Hoang, C. Grimaldier, P. Colson, et al., Viral RNA load as determined by cell culture as a management tool for discharge of SARS-CoV-2 patients from infectious disease wards, Eur. J. Clin. Microbiol. Infect. Dis., 39 (2020), 1059–1061. https://doi.org/10.1007/s10096-020-03913-9 doi: 10.1007/s10096-020-03913-9
|
| [47] | A. Singanayagam, M. Patel, A. Charlett, J. L. Bernal, V. Saliba, J. Ellis, et al., Duration of infectiousness and correlation with RT-PCR cycle threshold values in cases of COVID-19, England, January to May 2020, Eurosurveillance, 25 (2020), 2001483. |
| [48] |
A. Piralla, M. Ricchi, M. G. Cusi, P. Prati, N. Vicari, G. Scarsi, et al., Residual SARS-CoV-2 RNA in nasal swabs of convalescent COVID-19 patients: Is prolonged quarantine always justified?, IJID, 102 (2021), 299–302. https://doi.org/10.1016/j.ijid.2020.10.072 doi: 10.1016/j.ijid.2020.10.072
|
| [49] | M. J. Keeling, S. Moore, B. S. Penman, E. M. Hill, The impacts of SARS-CoV-2 vaccine dose separation and targeting on the COVID-19 epidemic in England, Nat. Commun., 14 (2023). https://doi.org/10.1038/s41467-023-35943-0 |
| [50] |
Y. Liu, L. M. Yan, L. Wan, T. X. Xiang, A. Le, J. M. Liu, et al., Viral dynamics in mild and severe cases of COVID-19, Lancet Infect. Dis., 20 (2020), 656–657. https://doi.org/10.1016/S1473-3099(20)30232-2 doi: 10.1016/S1473-3099(20)30232-2
|
| [51] |
A. Medaglia, G. Colelli, L. Farina, A. Bacila, P. Bini, E. Marchioni, et al., Uncertainty quantification and control of kinetic models of tumour growth under clinical uncertainties, Int. J. NonLin. Mech., 141 (2022), 103933. https://doi.org/10.1016/j.ijnonlinmec.2022.103933 doi: 10.1016/j.ijnonlinmec.2022.103933
|
Figures(6) / Tables(3)

Andrea Bondesan, Antonio Piralla, Elena Ballante, Antonino Maria Guglielmo Pitrolo, Silvia Figini, Fausto Baldanti, Mattia Zanella. Predictability of viral load dynamics in the early phases of SARS-CoV-2 through a model-based approach[J]. Mathematical Biosciences and Engineering, 2025, 22(4): 725-743. doi: 10.3934/mbe.2025027







 DownLoad:
DownLoad: