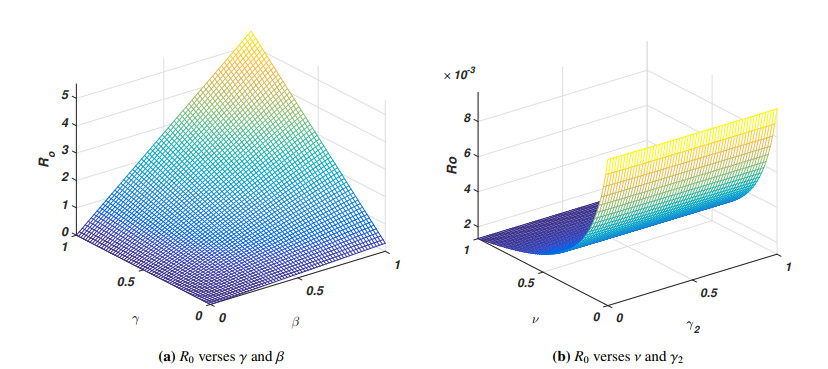
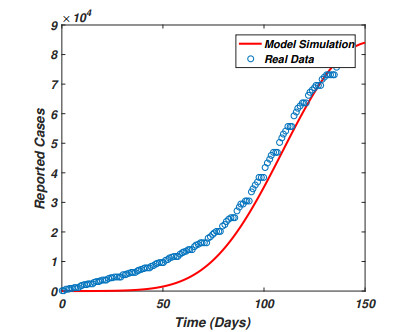
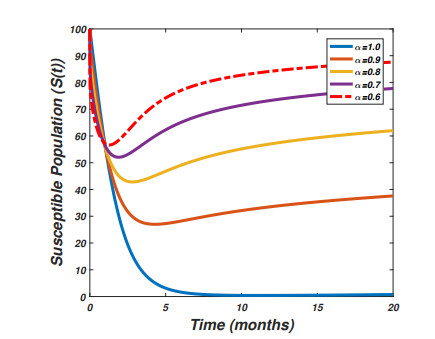
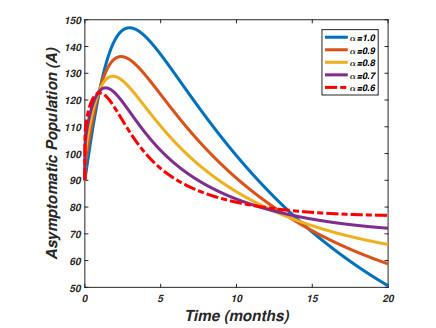
The pandemic of SARS-CoV-2 virus remains a pressing issue with unpredictable characteristics which spread worldwide through human interactions. The current study is focusing on the investigation and analysis of a fractional-order epidemic model that discusses the temporal dynamics of the SARS-CoV-2 virus in a community. It is well known that symptomatic and asymptomatic individuals have a major effect on the dynamics of the SARS-CoV-2 virus therefore, we divide the total population into susceptible, asymptomatic, symptomatic, and recovered groups of the population. Further, we assume that the vaccine confers permanent immunity because multiple vaccinations have commenced across the globe. The new fractional-order model for the transmission dynamics of SARS-CoV-2 virus is formulated via the Caputo-Fabrizio fractional-order approach with the maintenance of dimension during the process of fractionalization. The theory of fixed point will be used to show that the proposed model possesses a unique solution whereas the well-posedness (bounded-ness and positivity) of the fractional-order model solutions are discussed. The steady states of the model are analyzed and the sensitivity analysis of the basic reproductive number is explored. Moreover to parameterize the model a real data of SARS-CoV-2 virus reported in the Sultanate of Oman from January 1st, 2021 to May 23rd, 2021 are used. We then perform the large scale numerical findings to show the validity of the analytical work.
Citation: Tahir Khan, Roman Ullah, Gul Zaman, Jehad Alzabut. A mathematical model for the dynamics of SARS-CoV-2 virus using the Caputo-Fabrizio operator[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6095-6116. doi: 10.3934/mbe.2021305
The pandemic of SARS-CoV-2 virus remains a pressing issue with unpredictable characteristics which spread worldwide through human interactions. The current study is focusing on the investigation and analysis of a fractional-order epidemic model that discusses the temporal dynamics of the SARS-CoV-2 virus in a community. It is well known that symptomatic and asymptomatic individuals have a major effect on the dynamics of the SARS-CoV-2 virus therefore, we divide the total population into susceptible, asymptomatic, symptomatic, and recovered groups of the population. Further, we assume that the vaccine confers permanent immunity because multiple vaccinations have commenced across the globe. The new fractional-order model for the transmission dynamics of SARS-CoV-2 virus is formulated via the Caputo-Fabrizio fractional-order approach with the maintenance of dimension during the process of fractionalization. The theory of fixed point will be used to show that the proposed model possesses a unique solution whereas the well-posedness (bounded-ness and positivity) of the fractional-order model solutions are discussed. The steady states of the model are analyzed and the sensitivity analysis of the basic reproductive number is explored. Moreover to parameterize the model a real data of SARS-CoV-2 virus reported in the Sultanate of Oman from January 1st, 2021 to May 23rd, 2021 are used. We then perform the large scale numerical findings to show the validity of the analytical work.

| [1] | S. G. Samko, Fractional Integrals and Derivatives, Theory and Applications, Minsk, Nauka I Tekhnika, 1987. |
| [2] | I. Podlubny, Fractional Differential Equations: An Introduction to Fractional Derivatives, Fractional Differential Equations, to Methods of Their Solution and Some of Their Applications, Elsevier, 1999. |
| [3] | D. Baleanu, Z. B. Güvenç, J. T. Machado, New Trends in Nanotechnology and Fractional Calculus Applications, Springer, New York, 2010. |
| [4] | D. Baleanu, J. A. T. Machado, A. C. Luo, Fractional Dynamics and Control, Springer Science and Business Media, 2011. |
| [5] |
G. Zaman, Y. H. Kang, I. H. Jung, Stability analysis and optimal vaccination of an SIR epidemic model, BioSystems, 93 (2008), 240-249. doi: 10.1016/j.biosystems.2008.05.004

|
| [6] | Y. Wang, J. Cao, Global dynamics of a network epidemic model for waterborne diseases spread, Appl. Math. Comput., 237 (2014), 474–488. |
| [7] |
H. Abboubakar, J. C. Kamgang, D. Tieudjo, Backward bifurcation and control in transmission dynamics of arboviral diseases, Math. Biosci., 278 (2016), 100-129. doi: 10.1016/j.mbs.2016.06.002
|
| [8] |
T. Khan, G. Zaman, M. I. Chohan, The transmission dynamic and optimal control of acute and chronic hepatitis B, J. Biol. Dyn., 11 (2017), 172-189. doi: 10.1080/17513758.2016.1256441
|
| [9] | Y. Tu, S. Gao, Y. Liu, D. Chen, Y. Xu, Transmission dynamics and optimal control of stage-structured HLB model, J. Biol. Dyn., 16 (2019), 5180. |
| [10] |
A. Akgül, S. H. Khoshnaw, A. S. Abdalrahman, Mathematical modeling for enzyme inhibitors with slow and fast subsystems, Arab J. Basic Appl. Sci., 27 (2020), 442–449. doi: 10.1080/25765299.2020.1844369
|
| [11] |
A. Atangana, Modelling the spread of COVID-19 with new fractal-fractional operators: can the lockdown save mankind before vaccination, Chaos, Solitons Fractals, 136 (2020), 109860. doi: 10.1016/j.chaos.2020.109860
|
| [12] |
B. Ghanbari, A. Atangana, A new application of fractional Atangana–Baleanu derivatives: designing ABC-fractional masks in image processing, Physica A: Stat. Mech. Appl., 542 (2020), 123516. doi: 10.1016/j.physa.2019.123516
|
| [13] | A. G. M. Selvam, J. Alzabut, D. A. Vianny, M. Jacintha, F. B. Yousef, Modeling and stability analysis of the spread of novel coronavirus disease COVID-19, Int. J. Biomath., 2021 (2021), 2150035. |
| [14] |
F. Bozkurt, A. Yousef, D. Baleanu, J. Alzabut, A mathematical model of the evolution and spread of pathogenic coronaviruses from natural host to human host, Chaos, Solitons Fractals, 138 (2020), 109931. doi: 10.1016/j.chaos.2020.109931
|
| [15] |
K. M. Owolabi, A. Atangana, A. Akgul, Modelling and analysis of fractal-fractional partial differential equations: application to reaction-diffusion model, Alexandria Eng. J., 59 (2020), 2477–2490. doi: 10.1016/j.aej.2020.03.022
|
| [16] | H. Mohammadi, S. Rezapour, A. Jajarmi, On the fractional SIRD mathematical model and control for the transmission of COVID-19: the first and the second waves of the disease in Iran and Japan, ISA Trans., 2021. |
| [17] | A. A. Kilbas, H. M. Srivastava, J. J. Trujillo, Theory and Applications of Fractional Differential Equations, Elsevier, 204 (2006). |
| [18] |
H. M. Srivastava, K. M. Saad, J. F. Gómez-Aguilar, A. A. Almadiy, Some new mathematical models of the fractional-order system of human immune against IAV infection, Math. Biosci. Eng., 17 (2020), 4942–4969. doi: 10.3934/mbe.2020268
|
| [19] |
N. H. Tuan, H. Mohammadi, S. Rezapour, A mathematical model for COVID-19 transmission by using the Caputo fractional derivative, Chaos, Solitons Fractals, 140 (2020), 110107. doi: 10.1016/j.chaos.2020.110107
|
| [20] |
L. F. Ávalos-Ruiz, J. F. Gomez-Aguilar, A. Atangana, K. M. Owolabi, On the dynamics of fractional maps with power-law, exponential decay and Mittag–Leffler memory, Chaos, Solitons Fractals, 127 (2019), 364–388. doi: 10.1016/j.chaos.2019.07.010
|
| [21] |
K. M. Owolabi, A. Atangana, Numerical approximation of nonlinear fractional parabolic differential equations with Caputo–Fabrizio derivative in Riemann–Liouville sense, Chaos, Solitons Fractals, 99 (2017), 171–179. doi: 10.1016/j.chaos.2017.04.008
|
| [22] |
O. A. Arqub, Application of residual power series method for the solution of time-fractional Schrödinger equations in one-dimensional space, Fundam. Informaticae, 166 (2019), 87–110. doi: 10.3233/FI-2019-1795
|
| [23] |
O. A. Arqub, Numerical algorithm for the solutions of fractional order systems of Dirichlet function types with comparative analysis, Fundam. Informaticae, 166 (2019), 111–137. doi: 10.3233/FI-2019-1796
|
| [24] | S. Rezapour, H. Mohammadi, M. E. Samei, SEIR epidemic model for COVID-19 transmission by Caputo derivative of fractional order. Advances in difference equations, Adv. Differ. Equations, 1 (2020), 1–9. |
| [25] | M. Caputo, Michele, M. Fabrizio, A new definition of fractional derivative without singular kernel, Progr. Fract. Differ. Appl., 1 (2015), 1–13. |
| [26] |
H. A. A. El-Saka, The fractional-order SIS epidemic model with variable population size, J. Egypt. Math. Soc., 22 (2014), 50–54. doi: 10.1016/j.joems.2013.06.006
|
| [27] |
R. Toledo-Hernandez, V. Rico-Ramirez, G. A. Iglesias-Silva, U. M. Diwekar, A fractional calculus approach to the dynamic optimization of biological reactive systems. Part I: Fractional models for biological reactions, Chem. Eng. Sci., 117 (2014), 217–228. doi: 10.1016/j.ces.2014.06.034
|
| [28] |
K. M. Owolabi, Computational analysis of different Pseudoplatystoma species patterns the Caputo-Fabrizio derivative, Chaos, Solitons Fractals, 144 (2021), 110675. doi: 10.1016/j.chaos.2021.110675
|
| [29] |
D. Baleanu, A. Mousalou, S. Rezapour, A new method for investigating approximate solutions of some fractional integro-differential equations involving the Caputo-Fabrizio derivative, Adv. Differ. Equations, 2017 (2017), 1–12. doi: 10.1186/s13662-016-1057-2
|
| [30] |
M. S. Aydogan, D. Baleanu, A. Mousalou, S. Rezapour, On high order fractional integro-differential equations including the Caputo-Fabrizio derivative, Boundary Value Probl., 2018 (2018), 1–15. doi: 10.1186/s13661-017-0918-2
|
| [31] |
D. Baleanu, S. Rezapour, Z. Saberpour, On fractional integro-differential inclusions via the extended fractional Caputo-Fabrizio derivation, Boundary Value Probl., 2019 (2019), 1–17. doi: 10.1186/s13661-018-1115-7
|
| [32] |
D. Baleanu, A. Jajarmi, H. Mohammadi, S. Rezapour, A new study on the mathematical modelling of human liver with Caputo–Fabrizio fractional derivative, Chaos, Solitons Fractals, 134 (2020), 109705. doi: 10.1016/j.chaos.2020.109705
|
| [33] |
S. Rezapour, H. Mohammadi, A study on the AH1N1/09 influenza transmission model with the fractional Caputo-Fabrizio derivative, Adv. Differ. Equations, 2020 (2020), 1–15. doi: 10.1186/s13662-019-2438-0
|
| [34] |
S. Rezapour, H. Mohammadi, A. Jajarmi, A new mathematical model for Zika virus transmission, Adv. Differ. Equations, 2020 (2020), 1–15. doi: 10.1186/s13662-019-2438-0
|
| [35] |
D. Baleanu, H. Mohammadi, S. Rezapour, A mathematical theoretical study of a particular system of Caputo-Fabrizio fractional differential equations for the Rubella disease model, Adv. Differ. Equations, 2020 (2020), 1–19. doi: 10.1186/s13662-019-2438-0
|
| [36] | N. M. Ferguson, D. Laydon, G. Nedjati-Gilani, N. Imai, K. Ainslie, M. Baguelin, et al., Impact of non-pharmaceutical interventions (NPIs) to reduce COVID-19 mortality and healthcare demand. Imperial College COVID-19 Response Team, Imp. Coll. COVID-19 Response Team, 2020. |
| [37] |
A. J. Kucharski, T. W. Russell, C. Diamond, Y. Liu, J. Edmunds, S. Funk, et al., Early dynamics of transmission and control of COVID-19: a mathematical modelling study, Lancet Infect. Dis., 20 (2020), 553–558. doi: 10.1016/S1473-3099(20)30144-4
|
| [38] |
T. Kuniya, Prediction of the epidemic peak of coronavirus disease in Japan, 2020, J. Clin. Med., 9 (2020), 789. doi: 10.3390/jcm9030789
|
| [39] | R. O. Stutt, R. Retkute, M. Bradley, C. A. Gilligan, J. Colvin, A modelling framework to assess the likely effectiveness of facemasks in combination with 'lock-down'in managing the COVID-19 pandemic, Proc. R. Soc. A, 476 (2020), 2238. |
| [40] | Z. Tang, X. Li, H. Li, Prediction of new coronavirus infection based on a modified SEIR model, medRxiv, 2020. |
| [41] |
Z. Tang, X. Li, H. Li, New applications related to Covid-19, Results Phys., 20 (2021), 103663. doi: 10.1016/j.rinp.2020.103663
|
| [42] |
A. Atangana, S. İ. Araz, Nonlinear equations with global differential and integral operators: existence, uniqueness with application to epidemiology, Results Phys., 20 (2021), 103593. doi: 10.1016/j.rinp.2020.103593
|
| [43] | M. Rahman, S. Ahmad, R. T. Matoog, N. A. Alshehri, T. Khan, Study on the mathematical modelling of COVID-19 with Caputo-Fabrizio operator, Chaos, Solitons Fractals, 2021 (2021), 111121. |
| [44] |
H. M. Dobrovolny, Modeling the role of asymptomatics in infection spread with application to SARS-CoV-2, Plos one, 15 (2020), e0236976. doi: 10.1371/journal.pone.0236976
|
| [45] |
H. M. Dobrovolny, Quantifying the effect of remdesivir in rhesus macaques infected with SARS-CoV-2, Virology, 550 (2020), 61–69. doi: 10.1016/j.virol.2020.07.015
|
| [46] | S. Mandal, T. Bhatnagar, N. Arinaminpathy, A. Agarwal, A. Chowdhury, M. Murhekar, et al., Prudent public health intervention strategies to control the coronavirus disease 2019 transmission in India: A mathematical model-based approach, Indian J. Med. Res., 151 (2020), 2–3. |
| [47] |
R. F. Reis, B. de Melo Quintela, J. de Oliveira Campos, J. M. Gomes, B. M. Rocha, M. Lobosco, et al., Characterization of the COVID-19 pandemic and the impact of uncertainties, mitigation strategies, and underreporting of cases in South Korea, Italy, and Brazil, Chaos, Solitons Fractals, 136 (2020), 109888. doi: 10.1016/j.chaos.2020.109888
|
| [48] | X. Yang, L. Chen, J. Chen, Permanence and positive periodic solution for the single-species nonautonomous delay diffusive models, Comput. Math. Appl., 32 (1996), 109–116. |
| [49] |
H. Ramos, Z. Kalogiratou, T. Monovasilis, T. E. Simos, An optimized two-step hybrid block method for solving general second order initial-value problems, Numer. Algorithms, 72 (2016), 1089–1102. doi: 10.1007/s11075-015-0081-8
|
| [50] |
J. Vigo-Aguiar, H. Ramos, C. Clavero, A first approach in solving initial-value problems in ODEs by elliptic fitting methods, J. Comput. Appl. Math., 318 (2017), 599–603. doi: 10.1016/j.cam.2015.12.005
|
| [51] | C. Li, F. Zeng, Numerical Methods for Fractional Calculus, Chapman and Hall/CRC, 2019. |
| [52] |
A. Jajarmi, D. Baleanu, A new fractional analysis on the interaction of HIV with CD4+ T-cells, Chaos, Solitons Fractals, 113 (2018), 221–229. doi: 10.1016/j.chaos.2018.06.009
|
| [53] |
A. Jajarmi, D. Baleanu, On the nonlinear dynamical systems within the generalized fractional derivatives with Mittag-Leffler kernel, Nonlinear Dyn., 94 (2018), 397–414. doi: 10.1007/s11071-018-4367-y
|
Figures(6) / Tables(1)

Tahir Khan, Roman Ullah, Gul Zaman, Jehad Alzabut. A mathematical model for the dynamics of SARS-CoV-2 virus using the Caputo-Fabrizio operator[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 6095-6116. doi: 10.3934/mbe.2021305







 DownLoad:
DownLoad: