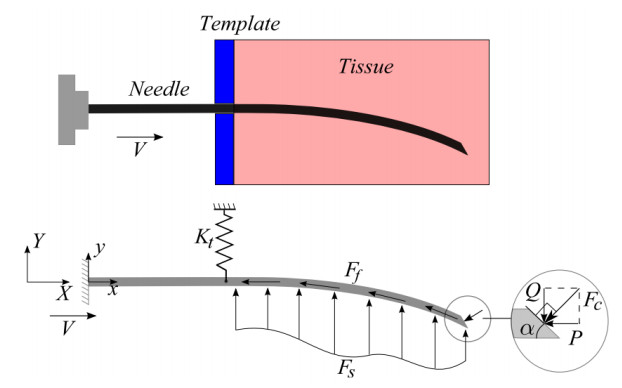
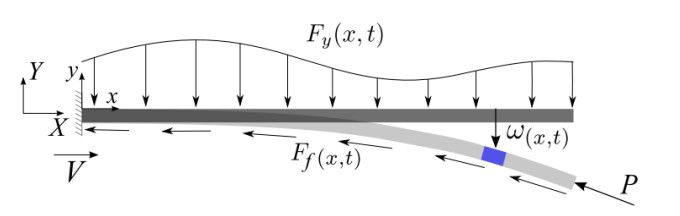
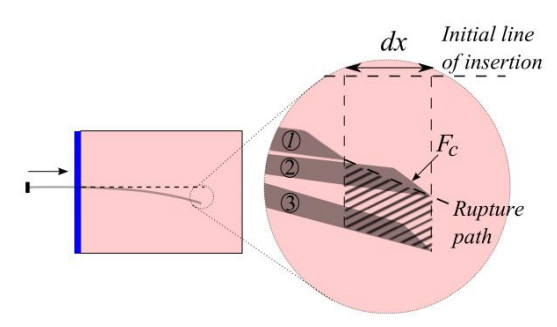
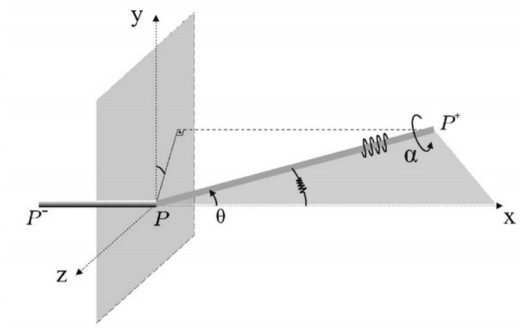
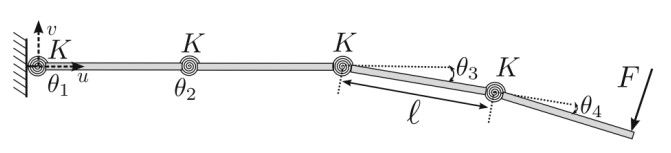
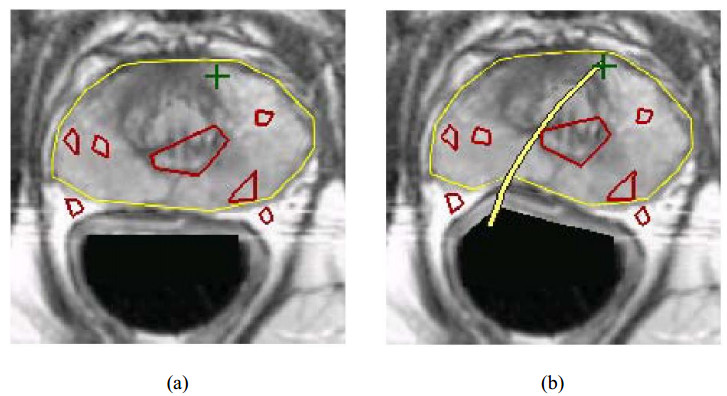
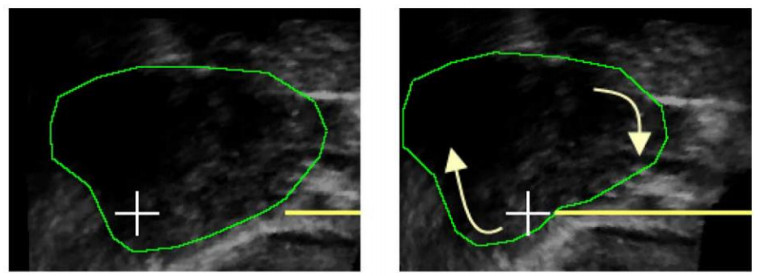
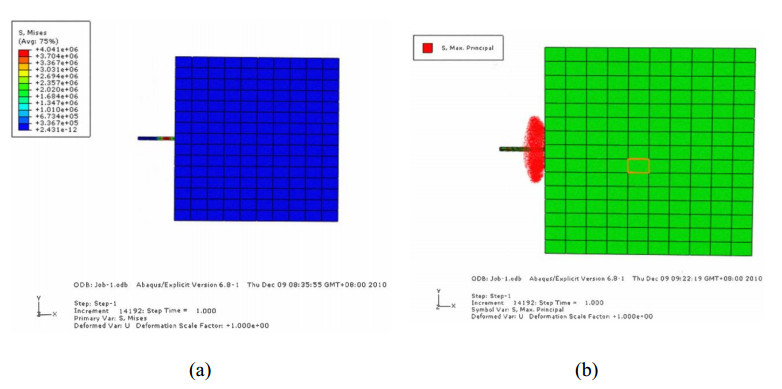
A flexible needle has emerged as a crucial clinical technique in contemporary medical practices, particularly for minimally invasive interventions. Its applicability spans diverse surgical domains such as brachytherapy, cardiovascular surgery, neurosurgery and others. Notably, flexible needles find utility in biopsies requiring deep skin penetration to access infected areas. Despite its minimally invasive advantages, the precise guidance of the needle to its intended target, while avoiding damage to bones, blood vessels, organs and tissues, remains a significant challenge for researchers. Consequently, extensive research has been dedicated to enhancing the steering and accuracy of flexible needles. Here, we aim to elucidate the recent advancements, trends and perspectives in flexible needle steering models and path planning over the last 15 years. The discussed models encompass various types, including symmetric-tip needles, curved-tip needles, tendon-actuated needles, programmable needles and the innovative fracture-directed waterjet needles. Moreover, the paper offers a comprehensive analysis, comparing the trajectories followed by these needle models to attain the desired target with minimal tissue damage. By delving into these aspects, the paper contributes to a deeper understanding of the current landscape of flexible needle technology and guides future research directions in this dynamic field.
Citation: Hafiz Muhammad Muzzammil, Yong-De Zhang, Hassan Ejaz, Qihang Yuan, Muhammad Muddassir. A review on tissue-needle interaction and path planning models for bevel tip type flexible needle minimal intervention[J]. Mathematical Biosciences and Engineering, 2024, 21(1): 523-561. doi: 10.3934/mbe.2024023
A flexible needle has emerged as a crucial clinical technique in contemporary medical practices, particularly for minimally invasive interventions. Its applicability spans diverse surgical domains such as brachytherapy, cardiovascular surgery, neurosurgery and others. Notably, flexible needles find utility in biopsies requiring deep skin penetration to access infected areas. Despite its minimally invasive advantages, the precise guidance of the needle to its intended target, while avoiding damage to bones, blood vessels, organs and tissues, remains a significant challenge for researchers. Consequently, extensive research has been dedicated to enhancing the steering and accuracy of flexible needles. Here, we aim to elucidate the recent advancements, trends and perspectives in flexible needle steering models and path planning over the last 15 years. The discussed models encompass various types, including symmetric-tip needles, curved-tip needles, tendon-actuated needles, programmable needles and the innovative fracture-directed waterjet needles. Moreover, the paper offers a comprehensive analysis, comparing the trajectories followed by these needle models to attain the desired target with minimal tissue damage. By delving into these aspects, the paper contributes to a deeper understanding of the current landscape of flexible needle technology and guides future research directions in this dynamic field.

| [1] |
R. Alterovitz, M. Branicky, K. Goldberg, Motion planning under uncertainty for image-guided medical needle steering, Int. J. Rob Res., 27 (2008), 1361–1374. https://doi.org/10.1177/0278364908097661 doi: 10.1177/0278364908097661

|
| [2] | Y. J. Zhao, Y. D. Zhang, F. Tu, Reverse path planning for flexible needle in 2D soft tissue with obstacles, Appl. Mech. Mater., 121-126 (2012), 4132–4137. https://doi.org/10.4028/www.scientific.net/AMM.121-126.4132 |
| [3] |
C. Rossa, N. Usmani, R. Sloboda, A hand-held assistant for semiautomated percutaneous needle steering, IEEE Trans. Biomed Eng., 64 (2017), 637–648. https://doi.org/10.1109/TBME.2016.2565690 doi: 10.1109/TBME.2016.2565690
|
| [4] |
C. Rossa, M Tavakoli, Issues in closed-loop needle steering, Control Eng. Pract., 62 (2017), 55–69. https://doi.org/10.1016/j.conengprac.2017.03.004 doi: 10.1016/j.conengprac.2017.03.004
|
| [5] | W. Park, J. S. Kim, Y. Zhou, Diffusion-based motion planning for a nonholonomic flexible needle model, in Proceedings of the IEEE Int. Conf. Robot Automation, (2005), 1050–4729. https://doi.org/10.1109/ROBOT.2005.1570829 |
| [6] |
R. J. Webster, J. S.Kin, N. J. Cowan, G. S. Chirikjian, A. M. Okamura, Nonholonomic modeling of needle steering, Tract.s Adv. Robot., 21 (2006), 35–44. https://doi.org/10.1177/0278364906065388 doi: 10.1177/0278364906065388
|
| [7] | S. Misra, K. T. Ramesh, A. M. Okamura, Modeling of tool-tissue interactions for computer-based surgical simulation: A lit review, Teleoper. Virt. Env., 17 (2008), 463–491. https://doi.org/10.1162/pres.17.5.463 |
| [8] |
D. Gao, Y. Lei, B. Lian, Modeling and Simulation of Flexible Needle Insertion into Soft Tissue Using Modified Local Constraints, J. Manuf. Sci. Eng. ASME, 138 (2016), 1–10. https://doi.org/10.1115/1.4034134 doi: 10.1115/1.4034134
|
| [9] |
T. D. Zhang, K. M. Shi, Y. J. Zhao, J. C. Yang, J. Liu, Path optimization algorithm and its robustness for bevel tip flexible needle, Int. J. Adv. Robot Syst., 15 (2018), 1–11. https://doi.org/10.1177/1729881418801166 doi: 10.1177/1729881418801166
|
| [10] |
D. Glozman, M. Shoham, Flexible needle steering and optimal trajectory planning for percutaneous therapies, Int. Conf. Med. Image Computer-assisted Intervent., 3217 (2004), 137–144. https://doi.org/10.1007/978-3-540-30136-3_18 doi: 10.1007/978-3-540-30136-3_18
|
| [11] | V. Duindam, R. Alterovitz, S. Sastry, K. Goldberg, Screw-based motion planning for bevel-tip flexible needles in 3D environments with obstacles, in Proc. - IEEE Int. Conf. Robot Autom., 22146911 (2008), 2483–2488. https://doi.org/10.1109%2FROBOT.2008.4543586 |
| [12] | R. Secoli, F. R. Y. Baena, Adaptive path-following control for bio-inspired steerable needles, in Proc. IEEE RAS EMBS Int. Conf. Biomed. Robot. Biomechatron., (2016), 87–93. https://doi.org/10.1109/BIOROB.2016.7523603 |
| [13] | R. Sun, T. Yang, Hybrid parameter-based PSO flexible needle percutaneous puncture path planning, J. Supercomput., (2023), 1–20. https://doi.org/10.1007/s11227-023-05661-x |
| [14] |
Z. Tan, D. Zhang, H. G. Liang, Q. G. Wang, W. Cai, A new path planning method for bevel-tip flexible needle insertion in 3D space with multiple targets and obstacles, Control Theory Technol., 20 (2022), 525–535. https://doi.org/10.1007/s11768-022-00113-y doi: 10.1007/s11768-022-00113-y
|
| [15] |
A. Segato, F. Calimeri, I. Testa, V. Corbetta, M. Riva, E. D. Momi, A hybrid inductive learning-based and deductive reasoning-based 3-D path planning method in complex environments, Auton. Robots, 46 (2022), 645–666. https://doi.org/10.1007/s10514-022-10042-z doi: 10.1007/s10514-022-10042-z
|
| [16] |
Z. Li, J. Dankelman, E. D. Momi, Path planning for endovascular catheterization under curvature constraints via two-phase searching approach, Int. J. Comput. Assist. Radiol. Surg., 16 (2021), 619–627. https://doi.org/10.1007/s11548-021-02328-x doi: 10.1007/s11548-021-02328-x
|
| [17] | J. Xue, V. Duindam, R. Alterovitz, K. Goldberg, Motion planning for steerable needles in 3D environments with obstacles using Rapidly-exploring random trees and backchaining, in 4th IEEE Conf. Autom. Sci. Eng. CASE, (2008), 41–46. https://doi.org/10.1109/COASE.2008.4626486 |
| [18] |
K. G. Yan, T. Podder, Y. Yu, T. Liu, C. W. Cheng, Flexible needle-tissue interaction modeling with depth-varying mean parameter: Preliminary study, IEEE Trans. Biomed. Eng., 56 (2009), 255–262. https://doi.org/10.1109/tbme.2008.2005959 doi: 10.1109/tbme.2008.2005959
|
| [19] |
O. Goksel, E. Dehghan, S. E. Salcudean, Modeling and simulation of flexible needles, Med. Eng. Phys., 31 (2009), 1069–1078. https://doi.org/10.1016/j.medengphy.2009.07.007 doi: 10.1016/j.medengphy.2009.07.007
|
| [20] | A. Asadian, M. R. Kermani, R. V. Patel, An analytical model for deflection of flexible needles during needle insertion, IEEE. Int. Conf. Intell. Robot. Syst., (2011), 2551–2556. https://doi.org/10.1109/IROS.2011.6094959 |
| [21] |
C. Rossa, R. Sloboda, N. Usmani, Estimating needle tip deflection in biological tissue from a single transverse ultrasound image: Application to brachytherapy, Int. J. Comput. Assist. Radiol. Surg., 11 (2016), 1347–1359. https://doi.org/10.1007/s11548-015-1329-4 doi: 10.1007/s11548-015-1329-4
|
| [22] | X. Zhao, L. Kong, D. Ye, Y. Zhao, Z. Wu, Flexible tip-steerable needle control with tissue uncertainty, in 26th Chinese Control Decis. Conf. CCDC, (2014), 4612–4616. https://doi.org/10.1109/CCDC.2014.6852996 |
| [23] |
M. Khadem, C. Rossa, N. Usmani, R. S. Sloboda, M. Tavakoli, Semi-Automated Needle Steering in Biological Tissue Using an Ultrasound-Based Deflection Predictor, Ann. Biomed. Eng., 45 (2017), 924–938. https://doi.org/10.1007/s10439-016-1736-x doi: 10.1007/s10439-016-1736-x
|
| [24] | M, Khadem, C. Rossa, N. Usmani, R. S. Sloboda, M. Tavakoli, Feedback-linearization-based 3D needle steering in a Frenet-Serret frame using a reduced order bicycle model, Proc. Am. Control Conf., (2017), 1438–1443. https://doi.org/10.23919/ACC.2017.7963155 |
| [25] |
M. Khadem, C. Rossa, N. Usmani, R. S. Sloboda, M. Tavakoli, Geometric control of 3D needle steering in soft-tissue, Automatica, 101 (2019), 36–43. https://doi.org/10.1016/j.automatica.2018.11.018 doi: 10.1016/j.automatica.2018.11.018
|
| [26] |
Y. Zhao, Z. Liu, Y. D. Zhang, Z. Q. Liu, Kinematic model and its parameter identification for cannula flexible needle insertion into soft tissue, Adv. Mech. Eng., 11 (2019), 1–13. https://doi.org/10.1177/1687814019852185 doi: 10.1177/1687814019852185
|
| [27] |
S. Misra, K. B. Reed, B. W. Schafer, K. T. Ramesh, A. M. Okamura, Mechanics of Flexible Needles Robotically Steered through Soft Tissue, Int. J. Rob. Res., 29 (2010), 1640–1660 https://doi.org/10.1177/0278364910369714 doi: 10.1177/0278364910369714
|
| [28] | R. J. Roesthuis, M. Abayazid, S. Misra, Mechanics-based model for predicting in-plane needle deflection with multiple bends, in Proc. IEEE RAS EMBS Int. Conf. Biomed. Robot Biomechatron., (2012), 69–74. https://doi.org/10.1109/BioRob.2012.6290829 |
| [29] | M. Khadem, B. Fallahi, C. Rossa, R. S. Sloboda, N. Usmani, M. Tavakoli, A mechanics-based model for simulation and control of flexible needle insertion in soft tissue, in Proc - IEEE Int. Conf. Robot. Autom., (2015), 2264–2269. https://doi.org/10.1109/ICRA.2015.7139499 |
| [30] |
C. Rossa, M. Khadem, R. Sloboda, N. Usmani, M. Tavakoli, Adaptive Quasi-Static Modelling of Needle Deflection during Steering in Soft Tissue, IEEE. Robot. Autom. Lett., 1 (2016), 916–923. https://doi.org/10.1109/LRA.2016.2527065 doi: 10.1109/LRA.2016.2527065
|
| [31] |
T. Lehmann, C. Rossa, N. Usmani, R. Sloboda, M. Tavakoli, Deflection modeling for a needle actuated by lateral force and axial rotation during insertion in soft phantom tissue, Mechatronics, 48 (2017), 42–53. https://doi.org/10.1016/j.mechatronics.2017.10.008 doi: 10.1016/j.mechatronics.2017.10.008
|
| [32] |
T. Watts, R. Secoli, F. R. Y. Baena, A Mechanics-Based Model for 3-D Steering of Programmable Bevel-Tip Needles, IEEE Trans. Robot., 35 (2019), 371–386. https://doi.org/10.1109/TRO.2018.2879584 doi: 10.1109/TRO.2018.2879584
|
| [33] |
D. Glozman, M. Shoham, Image-Guided Robotic Flexible Needle Steering, IEEE Trans. Robot., 23 (2007), 57–62. https://doi.org/10.1109/TRO.2007.898972 doi: 10.1109/TRO.2007.898972
|
| [34] | N. Sadati, M. Torabi, R. Vaziri, R. D. Ardekani, Soft-tissue modeling and image-guided control of steerable needles, in Proc. 31st Annu. Int. Conf. IEEE Eng. Med. Biol. Sci. Eng. Futur. Biomed. EMBC, (2009), 5122–5125. https://doi.org/10.1109/iembs.2009.5333473 |
| [35] |
W. park, Y. Liu, Y. Zhou, M. Moses, G. S. Chirikjian, Kinematic state estimation and motion planning for stochastic nonholonomic systems using the exponential map, Robotica, 26 (2008), 419–434. https://doi.org/10.1017/S0263574708004475 doi: 10.1017/S0263574708004475
|
| [36] |
R. Taschereau, J. Pouliot, J. Roy, D. Tremblay, Seed misplacement and stabilizing needles in transperineal permanent prostate implants, Radiother Oncol., 55 (2000), 59–63. https://doi.org/10.1016/s0167-8140(00)00162-6 doi: 10.1016/s0167-8140(00)00162-6
|
| [37] | R. Alterovitz, K. Goldberg, A. Okamura, Planning for steerable bevel-tip needle insertion through 2D soft tissue with obstacles, in Proc. - IEEE Int. Conf. Robot. Autom., (2005), 1640–1645. https://doi.org/10.1109/ROBOT.2005.1570348 |
| [38] |
R. Alterovitz, K. Y. Goldberg, J. Pouliot, I. J. Hsu, Sensorless motion planning for medical needle insertion in deformable tissues, IEEE. Trans. Inf. Technol. Biomed., 13 (2009), 217–225. https://doi.org/10.1109/titb.2008.2008393 doi: 10.1109/titb.2008.2008393
|
| [39] | Y. D. Zhang, F. Tu, The physical modeling of flexible needle biopsy soft tissue based on ABAQUS software, in 2011 IEEE/ICME Int. Conf. Complex. Med. Eng., (2011), 426–430. https://doi.org/10.1109/ICCME.2011.5876777 |
| [40] | S. Misra, K. B. Reed, A. S. Douglas, K. T. Ramesh, A. M. Okamura, Needle-tissue interaction forces for bevel-tip steerable needles, in Proc 2nd Bienn IEEE/RAS-EMBS Int, Conf, Biomed. Robot. Biomechatronics, (2008), 224–231. https://doi.org/10.1109%2FBIOROB.2008.4762872 |
| [41] |
M. C. Bernardes, B. V. Adorno, P. Poignet, G. A. Borges, Robot-assisted automatic insertion of steerable needles with closed-loop imaging feedback and intraoperative trajectory replanning, Mechatronics, 23 (2013), 630–645. https://doi.org/10.1016/j.mechatronics.2013.06.004 doi: 10.1016/j.mechatronics.2013.06.004
|
| [42] |
Y. J. Zhao, W. Q. Wu, Y. D. Zhang, R. X. Wang, J. C. Peng, Y. Yu, 3D dynamic motion planning for robot-assisted cannula flexible needle insertion into soft tissue, Int. J. Adv. Robot. Syst., 13 (2016), 1–11. https://doi.org/10.5772/64199 doi: 10.5772/64199
|
| [43] |
F. Liu, A. Garriga-Casanovas, R. Secoli, F. R. Y. Baena, Fast and adaptive fractal tree-based path planning for programmable bevel tip steerable needles, IEEE. Robot. Autom. Lett., 1 (2016), 601–608. https://doi.org/10.1109/LRA.2016.2528292 doi: 10.1109/LRA.2016.2528292
|
| [44] |
J. Van Den Berg, S. Patil, R. Alterovitz, P. Abbeel, K. Goldberg, LQG-based planning, sensing, and control of steerable needles, Springer Tracts. Adv. Robot., 68 (2010), 373–389. https://doi.org/10.1007/978-3-642-17452-0_22 doi: 10.1007/978-3-642-17452-0_22
|
| [45] | Y. J. Zhao, F. O. M. Joseph, K. Yan, N. V. Datla, Y. D. Zhang, T. K. Podder, et al. Path planning for robot-assisted active flexible needle using improved Rapidly-Exploring Random trees, in 2014 36th Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. EMBC, (2014), 380–383. https://doi.org/10.1109/embc.2014.6943608 |
| [46] |
P. Li, S. Jiang, D. Liang, Z. Yang, Y. Yu, W. Wang, Modeling of path planning and needle steering with path tracking in anatomical soft tissues for minimally invasive surgery, Med. Eng. Phys., 41 (2017), 35–45. https://doi.org/10.1016/j.medengphy.2017.01.006 doi: 10.1016/j.medengphy.2017.01.006
|
| [47] |
B. Fallahi, C. Rossa, R. S. Sloboda, N. Usmani, M. Tavakoli, Sliding-based image-guided 3D needle steering in soft tissue, Control. Eng. Pract., 63 (2017), 34–43. https://doi.org/10.1016/j.conengprac.2017.04.001 doi: 10.1016/j.conengprac.2017.04.001
|
| [48] |
W. Park, Y. Wang, G. S. Chirikjian, The path-of-probability algorithm for steering and feedback control of flexible needles, Int. J. Rob. Res., 29 (2010), 813–830. https://doi.org/10.1177/0278364909357228 doi: 10.1177/0278364909357228
|
| [49] | M. Lu, Y. Zhang, C. M. Lim, H. Ren, Flexible needle steering with tethered and untethered actuation: Current states, targeting errors, challenges and opportunities, Ann. Biomed. Eng., 51 (2023), 905–924. https://doi.org/10.1007/s10439-023-03163-8 |
| [50] |
M. Rox, M. Emerson, T. E. Ertop, I. Fried, M. Fu, J. Hoelscher, et al., Decoupling steerability from diameter: Helical dovetail laser patterning for steerable needles, IEEE Explore, 8 (2020), 181411–181419. https://doi.org/10.1109/ACCESS.2020.3028374 doi: 10.1109/ACCESS.2020.3028374
|
| [51] |
Z. K. Varnamkhasti, B. Konh, Compact 3D-printed active flexible needle for percutaneous procedures, Surg. Innov., 27 (2020), 402–405. https://doi.org/10.1177/1553350620945564 doi: 10.1177/1553350620945564
|
| [52] |
F. Yang, M. Babaiasl, J. P. Swensen, Fracture-Directed Steerable Needles, J. Med. Robot. Res., 4 (2019), 1842002. https://doi.org/10.1142/S2424905X18420023 doi: 10.1142/S2424905X18420023
|
| [53] |
S. Y. Ko, L. Frasson, Y. R. Y. Baena, Closed-loop planar motion control of a steerable probe with a programmable bevel inspired by nature, IEEE Trans. Robot., 27 (2011), 970–983. https://doi.org/10.1109/TRO.2011.2159411 doi: 10.1109/TRO.2011.2159411
|
| [54] | M. Scali, P. Breedveld, D. Dodou, Experimental evaluation of a self- propelling bio-inspired needle in single- and multi-layered phantoms, Sci. Rep., (2019), 1–13. https://doi.org/10.1038/s41598-019-56403-0 |
| [55] |
F. O. M. Joseph, T. Podder, Sliding mode control of a shape memory alloy actuated active flexible needle, Robotica, 36 (2018), 1188–1205. http://dx.doi.org/10.1017/S0263574718000334 doi: 10.1017/S0263574718000334
|
| [56] |
M. Khadem, C. Rossa, N. Usmani, R. S. Sloboda, M. Tavakoli, Robotic-Assisted Needle Steering Around Anatomical Obstacles Using Notched Steerable Needles, IEEE J. Biomed. Heal. Informatics, 22 (2017), 1917–1928. https://doi.org/10.1109/JBHI.2017.2780192 doi: 10.1109/JBHI.2017.2780192
|
| [57] | G. Gerboni, J. D. Greer, P. F. Laeseke, G. L. Hwang, A. M. Okamura, Highly articulated robotic needle achieves distributed ablation of liver tissue, IEEE Explore, 3766 (2017), 1–8. https://doi.org/10.1109%2FLRA.2017.2668467 |
| [58] |
A. Leibinger, M. J. Oldfield, F. R. Y. Baena, Minimally disruptive needle insertion: A biologically inspired solution, Interface Focus, 6 (2016), 1–10. https://doi.org/10.1098/rsfs.2015.0107 doi: 10.1098/rsfs.2015.0107
|
| [59] |
V. D. Berg, J. Dankelman, V. D. Dobbelsteen, Design of an actively controlled steerable needle with tendon actuation and FBG-based shape sensing, Med. Eng. Phys., 37 (2015), 617–622. https://doi.org/10.1016/j.medengphy.2015.03.016 doi: 10.1016/j.medengphy.2015.03.016
|
| [60] | R. Secoli, F. R. Y. Baena, Closed-loop 3D motion modeling and control of a steerable needle for soft tissue surgery, in Proc. - IEEE Int. Conf. Robot. Autom., (2013), 5831–5836. https://doi.org/10.1109/ICRA.2013.6631416 |
| [61] | L. Tang, Y. Chen, X He, Magnetic force aided compliant needle navigation and needle performance analysis, in 2007 IEEE Int. Conf. Robot. Biomimetics, (2007), 612–616. https://doi.org/10.1109/ROBIO.2007.4522232 |
| [62] | R. J. Webster, J. Memisevic, A. M. Okamura, Design considerations for needle steering, Proc. IEEE Int. Conf. Robot. Autom., (2005), 3599–3605. https://doi.org/10.1109/ROBOT.2005.1570666 |
| [63] |
C. Rossa, J. Fong, N. Usmani, R. Sloboda, M. Tavakoli, Multiactuator Haptic Feedback on the Wrist for Needle Steering Guidance in Brachytherapy, IEEE Robot. Autom. Lett., 1 (2016), 852–859. https://doi.org/10.1109/LRA.2016.2528295 doi: 10.1109/LRA.2016.2528295
|
| [64] |
J. Carriere, M. Khadem, C. Rossa, N. Usmani, R. Sloboda, M. Tavakoli, Event-Triggered 3D Needle Control Using a Reduced-Order Computationally Efficient Bicycle Model in a Constrained Optimization Framework, J. Med. Robotics. Res., 4 (2019), 1–16. https://doi.org/10.1142/S2424905X18420047 doi: 10.1142/S2424905X18420047
|
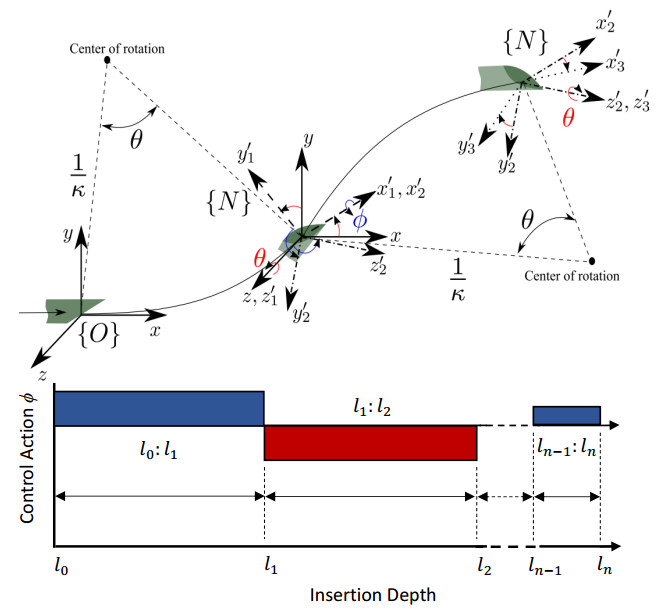
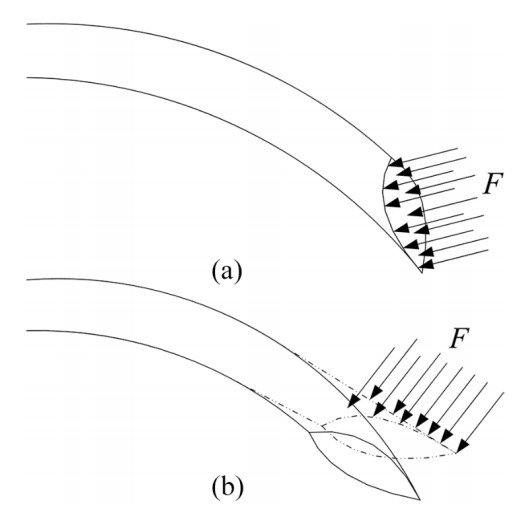
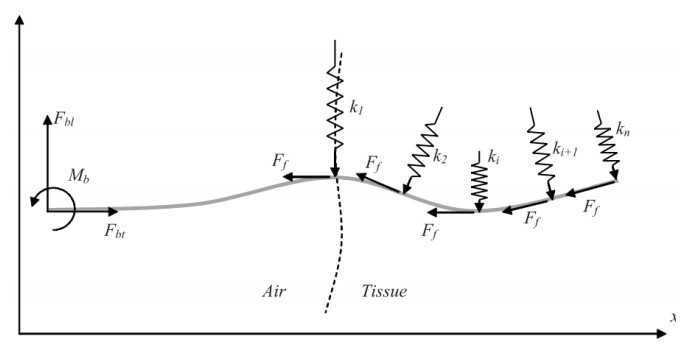
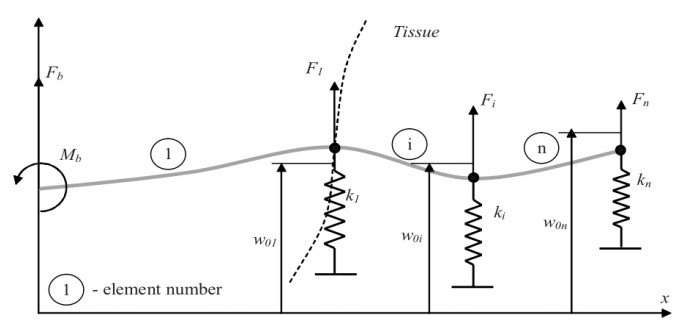
Figures(35) / Tables(4)

Hafiz Muhammad Muzzammil, Yong-De Zhang, Hassan Ejaz, Qihang Yuan, Muhammad Muddassir. A review on tissue-needle interaction and path planning models for bevel tip type flexible needle minimal intervention[J]. Mathematical Biosciences and Engineering, 2024, 21(1): 523-561. doi: 10.3934/mbe.2024023







 DownLoad:
DownLoad: