In this paper, dynamics analysis for a predator-prey model with strong Allee effect and nonconstant mortality rate are taken into account. We systematically studied the existence and stability of the equilibria, and detailedly analyzed various bifurcations, including transcritical, saddle-node, Hopf and Bogdanov-Takens bifurcation. In addition, the theoretical results are verified by numerical simulations. The results indicate that when the mortality is large, the nonconstant death rate can be approximated to a constant value. However, it cannot be considered constant under small mortality rate conditions. Unlike the extinction of species for the constant mortality, the nonconstant mortality may result in the coexistence of prey and predator for the predator-prey model with Allee effect.
Citation: Juan Ye, Yi Wang, Zhan Jin, Chuanjun Dai, Min Zhao. Dynamics of a predator-prey model with strong Allee effect and nonconstant mortality rate[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3402-3426. doi: 10.3934/mbe.2022157
In this paper, dynamics analysis for a predator-prey model with strong Allee effect and nonconstant mortality rate are taken into account. We systematically studied the existence and stability of the equilibria, and detailedly analyzed various bifurcations, including transcritical, saddle-node, Hopf and Bogdanov-Takens bifurcation. In addition, the theoretical results are verified by numerical simulations. The results indicate that when the mortality is large, the nonconstant death rate can be approximated to a constant value. However, it cannot be considered constant under small mortality rate conditions. Unlike the extinction of species for the constant mortality, the nonconstant mortality may result in the coexistence of prey and predator for the predator-prey model with Allee effect.

| [1] |
E. S$\acute{a}$ez, E. Gonz$\acute{a}$lez-Olivares, Dynamics of a predator-prey model, SIAM J. Appl. Math., 59 (1999), 1867–1878. https://doi.org/10.1137/S0036139997318457 doi: 10.1137/S0036139997318457

|
| [2] |
R. P. Gupta, M. Banerjee, P. Chandra, Bifurcation analysis and control of Leslie-Gower predator-prey model with Michaelis–Menten type prey-harvesting, Differ. Equations Dyn. Syst., 20 (2012), 339–366. https://doi.org/10.1007/s12591-012-0142-6 doi: 10.1007/s12591-012-0142-6
|
| [3] |
D. Hu, H. Cao, Stability and bifurcation analysis in a predator-prey system with Michaelis-Menten type predator harvesting, Nonlinear Anal.: Real World Appl., 33 (2017), 58–82. https://doi.org/10.1016/j.nonrwa.2016.05.010 doi: 10.1016/j.nonrwa.2016.05.010
|
| [4] |
L. Zhang, C. Zhang, M. Zhao, Dynamic complexities in a discrete predator–prey system with lower critical point for the prey, Math. Comput. Simul., 105 (2014), 119–131. https://doi.org/10.1016/j.matcom.2014.04.010 doi: 10.1016/j.matcom.2014.04.010
|
| [5] | C. Dai, M. Zhao, L. Chen, Dynamic complexity of an Ivlev-type prey-predator system with impulsive state feedback control, J. Appl. Math., 2012 (2012). https://doi.org/10.1155/2012/534276 |
| [6] |
C. Dai, M. Zhao, H. Yu, Dynamics induced by delay in a nutrient–phytoplankton model with diffusion, Ecol. Complexity, 26 (2016), 29–36. https://doi.org/10.1016/j.ecocom.2016.03.001 doi: 10.1016/j.ecocom.2016.03.001
|
| [7] |
C. Dai, M. Zhao, Mathematical and dynamic analysis of a prey-predator model in the presence of alternative prey with impulsive state feedback control, Discrete Dyn. Nat. Soc., 2012 (2012), 724014. https://doi.org/10.1155/2012/724014 doi: 10.1155/2012/724014
|
| [8] |
C. Dai, M. Zhao, Bifurcation and patterns induced by flow in a prey-predator system with Beddington-DeAngelis functional response, Phys. Rev. E, 102 (2020), 012209. https://doi.org/10.1103/PhysRevE.102.012209 doi: 10.1103/PhysRevE.102.012209
|
| [9] |
L. J. Wang, C. J. Dai, M. Zhao, Hopf bifurcation in an age-structured prey-predator model with Holling response function, Math. Biosci. Eng., 18 (2021), 3144–3159. https://doi.org/10.3934/mbe.2021156 doi: 10.3934/mbe.2021156
|
| [10] |
X. Y. Meng, Y. Q. Wu, Bifurcation analysis in a singular Beddington-DeAngelis predator-prey model with two delays and nonlinear predator harvesting, Math. Biosci. Eng., 16 (2019), 2668–2696. https://doi.org/10.3934/mbe.2019133 doi: 10.3934/mbe.2019133
|
| [11] |
P. Feng, On a diffusive predator-prey model with nonlinear harvesting, Math. Biosci. Eng., 11 (2014), 807. https://doi.org/10.3934/mbe.2014.11.807 doi: 10.3934/mbe.2014.11.807
|
| [12] |
P. A. Abrams, L. R. Ginzburg, The nature of predation: prey dependent, ratio dependent or neither, Trends Ecol. Evol., 15 (2000), 337–341. https://doi.org/10.1016/S0169-5347(00)01908-X doi: 10.1016/S0169-5347(00)01908-X
|
| [13] |
C. S. Holling, The components of predation as revealed by a study of small-mammal predation of the European Pine Sawfly1, Can. Entomol., 91 (1959), 293–320. https://doi.org/10.4039/Ent91293-5 doi: 10.4039/Ent91293-5
|
| [14] |
G. Seo, D. L. DeAngelis, A predator-prey model with a Holling type I functional response including a predator mutual interference, J. Nonlinear Sci., 21 (2011), 811–833. https://doi.org/10.1007/s00332-011-9101-6 doi: 10.1007/s00332-011-9101-6
|
| [15] |
W. Ko, K. Ryu, Qualitative analysis of a predator-prey model with Holling type II functional response incorporating a prey refuge, J. Differ. Equations, 231 (2006), 534–550. https://doi.org/10.1016/j.jde.2006.08.001 doi: 10.1016/j.jde.2006.08.001
|
| [16] |
E. Gonz$\acute{a}$lez-Olivares, A. Rojas-Palma, Multiple limit cycles in a Gause type predator–prey model with Holling type III functional response and Allee effect on prey, Bull. Math. Biol., 73 (2011), 1378–1397. https://doi.org/10.1007/s11538-010-9577-5 doi: 10.1007/s11538-010-9577-5
|
| [17] |
X. C. Zhang, G. Q. Sun, Z. Jin, Spatial dynamics in a predator-prey model with Beddington-DeAngelis functional response, Phys. Rev. E, 85 (2012), 021924. https://doi.org/10.1103/PhysRevE.85.021924 doi: 10.1103/PhysRevE.85.021924
|
| [18] |
D. Xiao, W. Li, M., Han, Dynamics in a ratio-dependent predator–prey model with predator harvesting, J. Math. Anal. Appl., 324 (2006), 14–29. https://doi.org/10.1016/j.jmaa.2005.11.048 doi: 10.1016/j.jmaa.2005.11.048
|
| [19] |
B. Liu, Y. Zhang, L. Chen, The dynamical behaviors of a Lotka–Volterra predator–prey model concerning integrated pest management, Nonlinear Anal.: Real World Appl., 6 (2005), 227–243. https://doi.org/10.1016/j.nonrwa.2004.08.001 doi: 10.1016/j.nonrwa.2004.08.001
|
| [20] |
M. Cavani, M. Farkas, Bifurcations in a predator-prey model with memory and diffusion. I: Andronov-Hopf bifurcation, Acta Math. Hung., 63 (1994), 213–229. https://doi.org/10.1007/BF01874129 doi: 10.1007/BF01874129
|
| [21] |
R. Arditi, L. R. Ginzburg, Coupling in predator-prey dynamics: ratio-dependence, J. Theor. Biol., 139 (1989), 311–326. https://doi.org/10.1016/S0022-5193(89)80211-5 doi: 10.1016/S0022-5193(89)80211-5
|
| [22] |
S. B. Hsu, S. P. Hubbell, P. Waltman, Competing predators, SIAM J. Appl. Math., 35 (1978), 617–625. https://doi.org/10.1137/0135051 doi: 10.1137/0135051
|
| [23] |
C. Dai, H. Liu, Z. Jin, Q. Guo, Y. Wang, H. Yu, et al., Dynamic analysis of a heterogeneous diffusive prey-predator system in time-periodic environment, Complexity, 2020 (2020), 1–13. https://doi.org/10.1155/2020/7134869 doi: 10.1155/2020/7134869
|
| [24] |
W. S. Yang, Dynamics of a diffusive predator-prey model with general nonlinear functional response, Sci. World J., 2014 (2014), 1–10. https://doi.org/10.1155/2014/721403 doi: 10.1155/2014/721403
|
| [25] |
C. Duque, M. Lizana, Partial characterization of the global dynamic of a predator-prey model with non constant mortality rate, Differ. Equations Dyn. Syst., 17 (2009), 63–75. https://doi.org/10.1007/s12591-009-0005-y doi: 10.1007/s12591-009-0005-y
|
| [26] |
C. Duque, M. Lizana, On the dynamics of a predator-prey model with nonconstant death rate and diffusion, Nonlinear Anal.: Real World Appl., 12 (2011), 2198–2210. https://doi.org/10.1016/j.nonrwa.2011.01.002 doi: 10.1016/j.nonrwa.2011.01.002
|
| [27] |
R. Yang, Hopf bifurcation analysis of a delayed diffusive predator-prey system with nonconstant death rate, Chaos, Solitons Fractals, 81 (2015), 224–232. https://doi.org/10.1016/j.chaos.2015.09.021 doi: 10.1016/j.chaos.2015.09.021
|
| [28] |
X. Fauvergue, J. C. Malausa, L. Giuge, F. Courchamp, Invading parasitoids suffer no Allee effect: a manipulative field experiment, Ecology, 88 (2007), 2392–2403. https://doi.org/10.1890/06-1238.1 doi: 10.1890/06-1238.1
|
| [29] |
A. M. Kramer, O. Sarnelle, R. A. Knapp, Allee effect limits colonization success of sexually reproducing zooplankton, Ecology, 89 (2008), 2760–2769. https://doi.org/10.1890/07-1505.1 doi: 10.1890/07-1505.1
|
| [30] |
Y. Huang, Z. Zhu, Z. Li, Modeling the Allee effect and fear effect in predator-prey system incorporating a prey refuge, Adv. Differ. Equations, 2020 (2020), 1–13. https://doi.org/10.1186/s13662-020-02727-5 doi: 10.1186/s13662-020-02727-5
|
| [31] |
D. Sen, S. Ghorai, S. Sharma, M. Banerjee, Allee effect in prey's growth reduces the dynamical complexity in prey-predator model with generalist predator, Appl. Math. Modell., 91 (2021), 768–790. https://doi.org/10.1016/j.apm.2020.09.046 doi: 10.1016/j.apm.2020.09.046
|
| [32] |
A. Kumar, B. Dubey, Dynamics of prey-predator model with strong and weak Allee effect in the prey with gestation delay, Nonlinear Anal.: Modell. Control, 25 (2020), 417–442. https://doi.org/10.15388/namc.2020.25.16663 doi: 10.15388/namc.2020.25.16663
|
| [33] |
Y. Kang, O. Udiani, Dynamics of a single species evolutionary model with Allee effects, J. Math. Anal. Appl., 418 (2014), 492–515. https://doi.org/10.1016/j.jmaa.2014.03.083 doi: 10.1016/j.jmaa.2014.03.083
|
| [34] |
X. Yu, S. Yuan, T. Zhang, Persistence and ergodicity of a stochastic single species model with Allee effect under regime switching, Commun. Nonlinear Sci. Numer. Simul., 59 (2018), 359–374. https://doi.org/10.1016/j.cnsns.2017.11.028 doi: 10.1016/j.cnsns.2017.11.028
|
| [35] |
E. Gonz$\acute{a}$lez-Olivares, J. Mena-Lorca, A. Rojas-Palma, J. D. Flores, Dynamical complexities in the Leslie–Gower predator–prey model as consequences of the Allee effect on prey, Appl. Math. Modell., 35 (2011), 366–381. https://doi.org/10.1016/j.apm.2010.07.001 doi: 10.1016/j.apm.2010.07.001
|
| [36] |
D. Mukherjee, Study of refuge use on a predator-prey system with a competitor for the prey, Int. J. Biomath., 10 (2017), 1750023. https://doi.org/10.1142/S1793524517500231 doi: 10.1142/S1793524517500231
|
| [37] |
R. K. Naji, S. J. Majeed, The dynamical analysis of a prey-predator model with a refuge-stage structure prey population, Int. J. Differ. Equations, 2016 (2016), 1–10. https://doi.org/10.1155/2016/2010464 doi: 10.1155/2016/2010464
|
| [38] |
Y. Zhang, X. Rong, J. Zhang, A diffusive predator-prey system with prey refuge and predator cannibalism, Math. Biosci. Eng., 16 (2019), 1445–1470. https://doi.org/10.3934/mbe.2019070 doi: 10.3934/mbe.2019070
|
| [39] |
F. Chen, L. Chen, X. Xie, On a Leslie–Gower predator–prey model incorporating a prey refuge, Nonlinear Anal.: Real World Appl., 10 (2009), 2905–2908. https://doi.org/10.1016/j.nonrwa.2008.09.009 doi: 10.1016/j.nonrwa.2008.09.009
|
| [40] |
L. Chen, F. Chen, L. Chen, Qualitative analysis of a predator–prey model with Holling type II functional response incorporating a constant prey refuge, Nonlinear Anal.: Real World Appl., 11 (2010), 246–252. https://doi.org/10.1016/j.nonrwa.2008.10.056 doi: 10.1016/j.nonrwa.2008.10.056
|
| [41] | Z. F. Zhang, T. R. Ding, W. Z. Huang, Z. X. Dong, Qualitative theory of differential equation, in Translations of Mathematical Monographs, 1992. https://doi.org/10.1090/mmono/101 |
| [42] |
D. Y. Wu, H. Y. Zhao, Y. Yuan, Complex dynamics of a diffusive predator-prey model with strong Allee effect and threshold harvesting, J. Math. Anal. Appl., 469 (2019), 982–1014. https://doi.org/10.1016/j.jmaa.2018.09.047 doi: 10.1016/j.jmaa.2018.09.047
|
| [43] |
D. Y. Wu, H. Y. Zhao, Spatiotemporal dynamics of a diffusive predator-prey system with Allee effect and threshold hunting, J. Nonlinear Sci., 30 (2020), 1015–1054. https://doi.org/10.1007/s00332-019-09600-0 doi: 10.1007/s00332-019-09600-0
|
| [44] | L. Perko, Differential Equations and Dynamical Systems, Springer, 2001. https://doi.org/10.1007/978-1-4613-0003-8 |
| [45] | J. Sotomayor, Generic bifurcations of dynamical systems, Dyn. Syst., 1973. https://doi.org/10.1016/B978-0-12-550350-1.50047-3 |
| [46] |
X. C. Zhang, G. Q. Sun, Z. Jin, Spatial dynamics in a predator-prey model with Beddington-DeAngelis functional response, Phys. Rev. E, 85 (2012), 021924. https://doi.org/10.1103/PhysRevE.85.021924 doi: 10.1103/PhysRevE.85.021924
|
| [47] |
X. B. Zhang, H. Y. Zhao, Stability and bifurcation of a reaction-diffusion predator-prey model with non-local delay and Michaelis-Menten-type prey-harvesting, Int. J. Comput. Math., 93 (2016), 1447–1469. https://doi.org/10.1080/00207160.2015.1056169 doi: 10.1080/00207160.2015.1056169
|
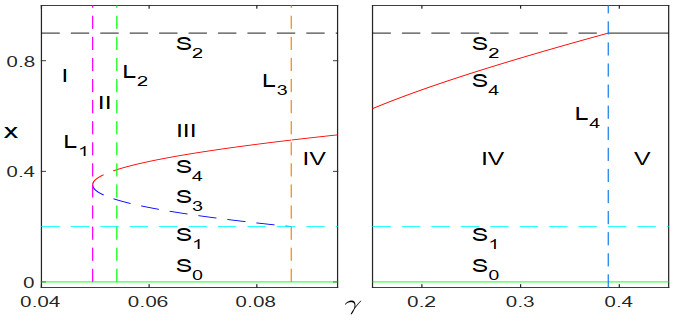
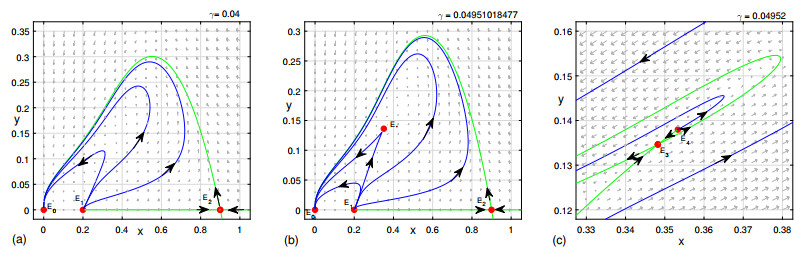
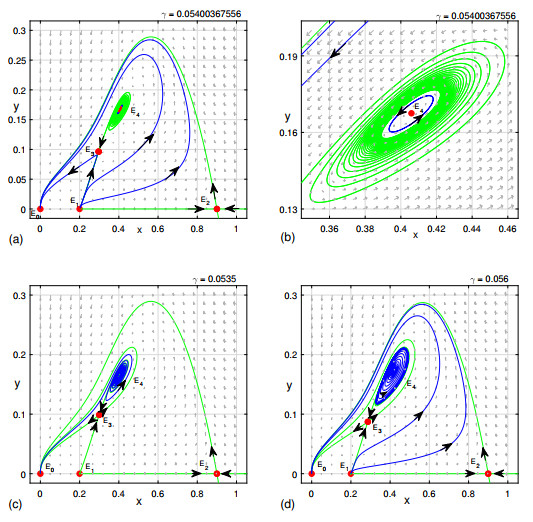
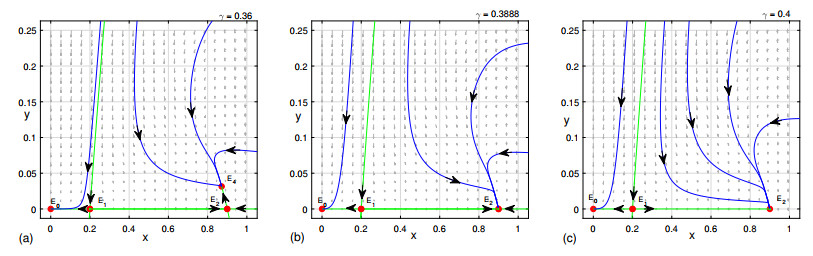
Figures(9)

Juan Ye, Yi Wang, Zhan Jin, Chuanjun Dai, Min Zhao. Dynamics of a predator-prey model with strong Allee effect and nonconstant mortality rate[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3402-3426. doi: 10.3934/mbe.2022157







 DownLoad:
DownLoad: