Citation: Bo Zheng, Lihong Chen, Qiwen Sun. Analyzing the control of dengue by releasing Wolbachia-infected male mosquitoes through a delay differential equation model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5531-5550. doi: 10.3934/mbe.2019275

| [1] | S. Bhatt, P. Gething, O. Brady, et al., The global distribution and burden of dengue, Nature, 496 (2013), 504–507. |
| [2] | Y. Wang, X. Liu, C. Li, et al., A survey of insecticide resistance in Aedes albopictus (Diptera: Culicidae) during a 2014 dengue fever outbreak in Guangzhou, China, J. Econ. Entomol., 110 (2017), 239–244. |
| [3] | F. Baldacchino, B. Caputo, F. Chandre, et al., Control methods against invasive Aedes mosquitoes in Europe: a review, Pest. Manag. Sci., 71 (2015), 1471–1485. |
| [4] | J. Werren, Biology of Wolbachia, Ann. Rev. Entomol., 42 (1997), 587–609. |
| [5] | G. Bian, D. Joshi, Y. Dong, et al., Wolbachia invades Anopheles stephensi populations and induces refractoriness to plasmodium infection, Science, 340 (2013), 748–751. |
| [6] | Z. Xi, C. Khoo and S. Dobson, Wolbachia establishment and invasion in an Aedes aegypti labora-tory population, Science, 310 (2005), 326–328. |
| [7] | E. Waltz, US reviews plan to infect mosquitoes with bacteria to stop disease, Nature, 89 (2016), 450–451. |
| [8] | M. Keeling, F. Jiggins and J. Read, The invasion and coexistence of competing Wolbachia strains, Heredity, 91 (2003), 382–388. |
| [9] | B. Zheng, M. Tang and J. Yu, Modeling Wolbachia spread in mosquitoes through delay differential equations, SIAM J. Appl. Math., 74 (2014), 743–770. |
| [10] | B. Zheng, M. Tang, J. Yu, et al., Wolbachia spreading dynamics in mosquitoes with imperfect maternal transmission, J. Math. Biol., 76 (2018), 235–263. |
| [11] | L. Cai, S. Ai and J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM J. Appl. Math., 74 (2014), 1786–1809. |
| [12] | L. Cai, S. Ai and G. Fan, Dynamics of delayed mosquitoes populations models with two different strategies of releasing sterile mosquitoes, Math. Biosci. Eng., 15 (2018), 1181–1202. |
| [13] | M. Huang, J. Luo, L. Hu, et al., Assessing the efficiency of Wolbachia driven Aedes mosquito suppression by delay differential equations, J. Theor. Biol., 440 (2018), 1–11. |
| [14] | Y. Li and X. Liu, An impulsive model for Wolbachia infection control of mosquito-borne diseases with general birth and death rate functions, Nonlinear Anal: RWA, 37 (2017), 412–432. |
| [15] | L. Hu, M. Huang, M. Tang, et al., Wolbachia spread dynamics in stochastic environments, Theor. Popul. Biol., 106 (2015), 32–44. |
| [16] | M. Turelli and N. Barton, Deploying dengue-suppressing Wolbachia: Robust models predict slow but effective spatial spread in Aedes aegypti, Theor. Popul. Biol., 115 (2017), 45–60. |
| [17] | E. Newton and P. Reiter, A model of the transmission of dengue fever with an evaluation of the impact of ultra-low volume (ULV) insecticide applications on dengue epidemics, Am. J. Trop. Med. Hyg., 47 (1992), 709–720. |
| [18] | M. Atkinson, Z. Su, N. Alphey, et al., Analyzing the control of mosquito-borne diseases by a dominant lethal genetic system, Proc. Natl. Acad. Sci. USA, 104 (2007), 9540–9545. |
| [19] | S. Pinho, C. Ferreira, L. Esteva, et al., Modelling the dynamics of dengue real epidemics, Phil. Trans. R. Soc. A, 368 (2010), 5679–5693. |
| [20] | L. Xue, X. Fang and J. Hyman, Comparing the effectiveness of different strains of Wolbachia for controlling chikungunya, dengue fever, and zika, PLoS Neglect. Trop. D, 12 (2018), e0006666. |
| [21] | J. Zhang, C. Yang, Z. Jin, et al., Dynamics analysis of SIR epidemic model with correlation coef-ficients and clustering coefficient in networks, J. Theor. Biol., 449 (2018), 1–13. |
| [22] | L. Zou, J. Chen, X. Feng, et al., Analysis of a dengue model with vertical transmission and ap-plication to the 2014 dengue outbreak in Guangdong Province, China, B. Math. Biol., 80 (2018), 2633–2651. |
| [23] | C. Craig, The Etiology of Dengue Fever, J. Am. Med. Assoc., 75 (1920), 1171–1176. |
| [24] | C. Dye, Models for the population dynamics of the yellow fever mosquito, Aedes aegypti, J. Anim. Ecol., 53 (1984), 247–268. |
| [25] | F. Coutinhoa, M. Burattinia, L. Lopeza, et al., Threshold conditions for a non-autonomous epi-demic system describing the population dynamics of Dengue, Bull. Math. Biol., 68 (2006), 2263–2282. |
| [26] | M. Guzman and E. Harris, Dengue, Lancet, 385 (2015), 453–465. |
| [27] | M. Chan and M. Johansson, The incubation periods of dengue viruses, PLoS One, 7 (2012), e50972. |
| [28] | H. Smith, An Introduction to Delay Differential Equations with Applications to the Life Sciences, Volume 57, Springer, New York, 2011. |
| [29] | National Bureau of Statistics of China, National Data, Accessed 20 Jan 2019. Available from: http://data.stats.gov.cn/. |
| [30] | T. Scott, A. Morrison, L. Lorenz, et al., Longitudinal studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: population dynamics, J. Med. Entomol., 37 (2000), 77–88. |
| [31] | C. Paupy, B. Ollomo, B. Kamgang, et al., Comparative role of Aedes albopictus and Aedes ae-gypti in the emergence of Dengue and Chikungunya in central Africa, Vector-Borne and Zoonotic Diseases, 10 (2010), 259–266. |
| [32] | Y. Ye, S. Chenoweth, A. Carrasco, et al., Evolutionary potential of the extrinsic incubation period of dengue virus in Aedes aegypti, Evolution, 70 (2016), 2459–2469. |
| [33] | J. Yu, Modeling mosquitoes population suppression based on delay differential equations, SIAM J. Appl. Math., 78 (2018), 3168–3187. |
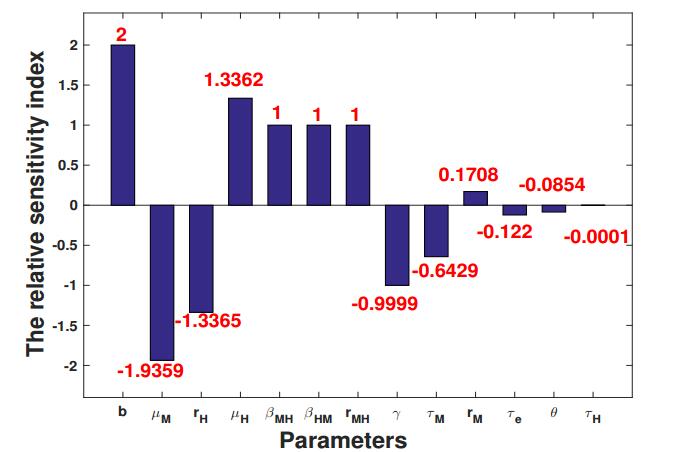
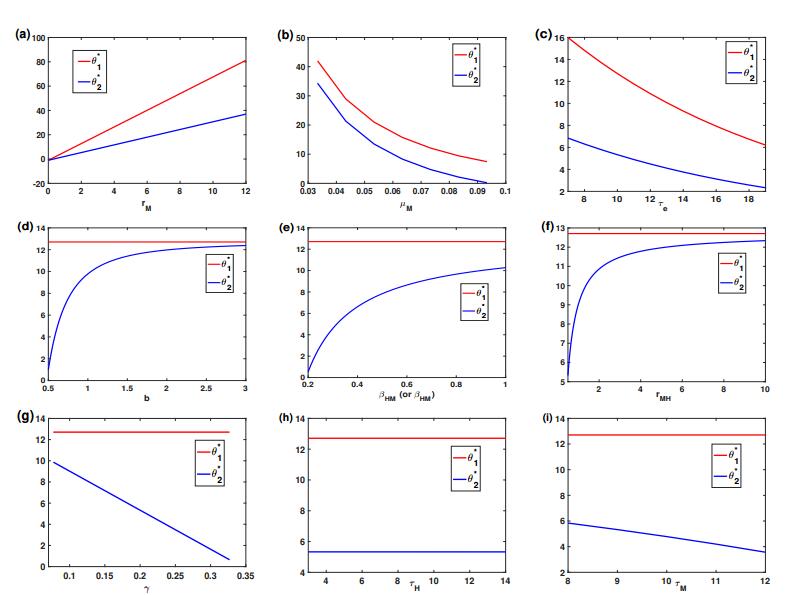
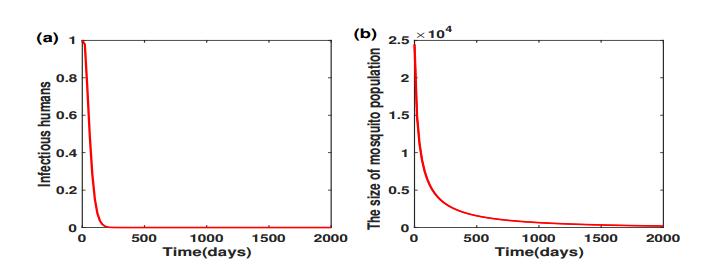
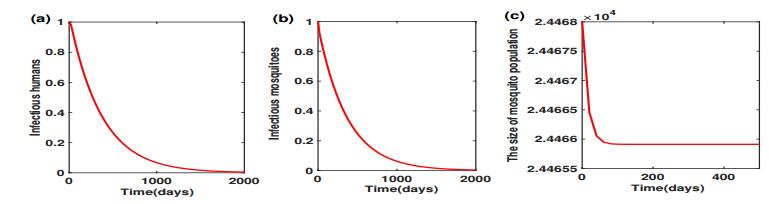
Figures(5) / Tables(2)

Bo Zheng, Lihong Chen, Qiwen Sun. Analyzing the control of dengue by releasing Wolbachia-infected male mosquitoes through a delay differential equation model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5531-5550. doi: 10.3934/mbe.2019275







 DownLoad:
DownLoad: