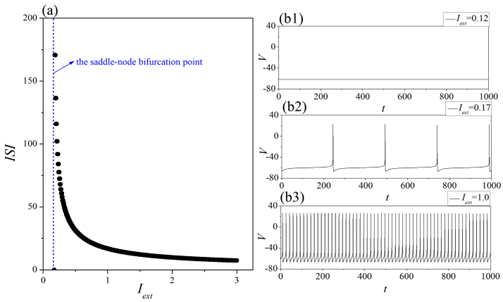
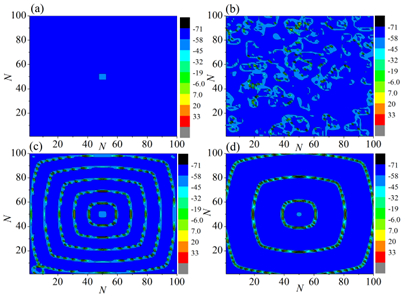
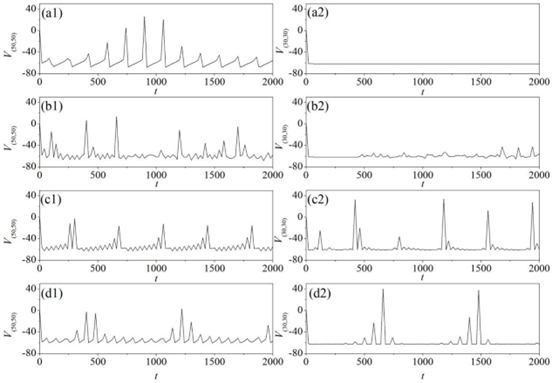
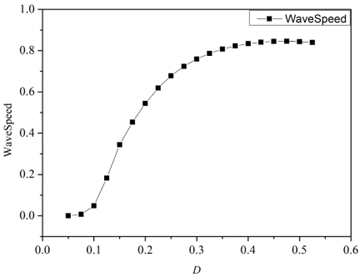
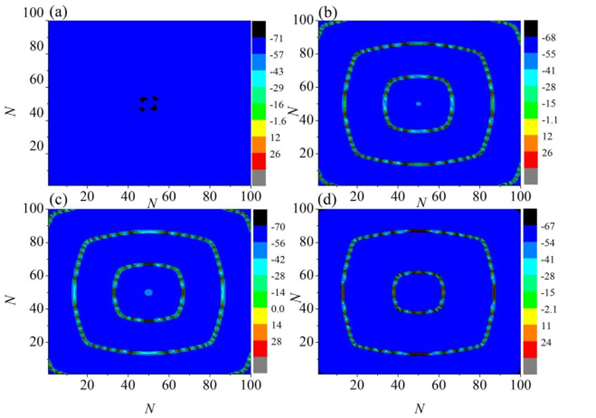
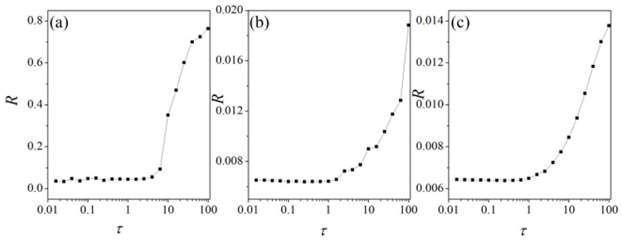
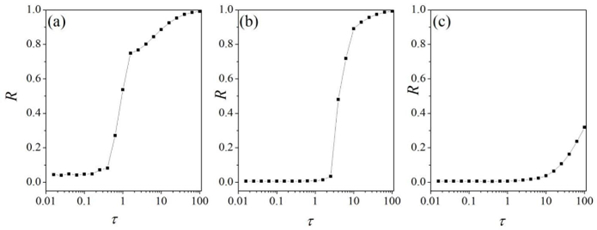
Interneurons modulate the excitability of neural networks and maintain neural activity balance via inhibitory or excitatory synaptic connections. Here, we studied the formation of patterns of interneuronal networks with inhibitory synaptic coupling. We found that both electrical synaptic coupling and inhibitory synaptic coupling play a crucial role in the formation of neural network patterns. In addition, delayed inhibitory synapses can also affect the transition of target waves to chaotic states. As the strength of electrical synaptic coupling increases, the firing behavior of neurons gradually becomes highly ordered. When the inhibitory synaptic delay reaches a critical value, we observe a transition in oscillatory patterns from an ordered state to a synchronized state. We further investigated how inhibitory synaptic conductance influences the formation of oscillatory patterns in the network. The study reveals that increasing synaptic conductance disrupts the structure of target waves, inducing chaotic states such as spiral wave fragmentation, while simultaneously elevating neuronal firing rates.
Citation: Ying Xu, Xiaodi Li. The pattern dynamics of interneuronal networks with inhibitory synaptic coupling[J]. AIMS Mathematics, 2025, 10(5): 10976-10993. doi: 10.3934/math.2025498
Interneurons modulate the excitability of neural networks and maintain neural activity balance via inhibitory or excitatory synaptic connections. Here, we studied the formation of patterns of interneuronal networks with inhibitory synaptic coupling. We found that both electrical synaptic coupling and inhibitory synaptic coupling play a crucial role in the formation of neural network patterns. In addition, delayed inhibitory synapses can also affect the transition of target waves to chaotic states. As the strength of electrical synaptic coupling increases, the firing behavior of neurons gradually becomes highly ordered. When the inhibitory synaptic delay reaches a critical value, we observe a transition in oscillatory patterns from an ordered state to a synchronized state. We further investigated how inhibitory synaptic conductance influences the formation of oscillatory patterns in the network. The study reveals that increasing synaptic conductance disrupts the structure of target waves, inducing chaotic states such as spiral wave fragmentation, while simultaneously elevating neuronal firing rates.

| [1] |
Y. Qian, X. Huang, G. Hu, X. Liao, Structure and control of self-sustained target waves in excitable small-world networks, Phys. Rev. E, 81 (2010), 036101. https://doi.org/10.1103/PhysRevE.81.036101 doi: 10.1103/PhysRevE.81.036101

|
| [2] |
Z. Rostami, V. T. Pham, S. Jafari, F. Hadaeghi, J. Ma, Taking control of initiated propagating wave in a neuronal network using magnetic radiation, Appl. Math. Comput., 338 (2018), 141–151. https://doi.org/10.1016/j.amc.2018.06.004 doi: 10.1016/j.amc.2018.06.004
|
| [3] |
C. Wang, M. Lv, A. Alsaedi, J. Ma, Synchronization stability and pattern selection in a memristive neuronal network, Chaos, 27 (2017), 113108. https://doi.org/10.1063/1.5004234 doi: 10.1063/1.5004234
|
| [4] |
H. Qin, C. Wang, N. Cai, X. An, F. Alzahrani, Field coupling-induced pattern formation in two-layer neuronal network, Physica A, 501 (2018), 141–152. https://doi.org/10.1016/j.physa.2018.02.063 doi: 10.1016/j.physa.2018.02.063
|
| [5] |
J. Ma, Y. Xu, G. Ren, C. Wang, Prediction for breakup of spiral wave in a regular neuronal network, Nonlinear Dyn., 84 (2016), 497–509. https://doi.org/10.1007/s11071-015-2502-6 doi: 10.1007/s11071-015-2502-6
|
| [6] |
Y. Wu, Q. Ding, W. Huang, T. Li, D. Yu, Y. Jia, Dynamic learning of synchronization in coupled nonlinear systems, Nonlinear Dyn., 112 (2024), 21945–21967. https://doi.org/10.1007/s11071-024-10192-y doi: 10.1007/s11071-024-10192-y
|
| [7] |
J. Ma, J. Tang, A review for dynamics in neuron and neuronal network, Nonlinear Dyn, 89 (2017), 1569–1578. https://doi.org/10.1007/s11071-017-3565-3 doi: 10.1007/s11071-017-3565-3
|
| [8] | Q. Ding, Y. Wu, W. Huang, Y. Jia, A dynamic learning method for phase synchronization control in Hodgkin–Huxley neuronal networks, Eur. Phys. J. Spec. Top., (2024). https://doi.org/10.1140/epjs/s11734-024-01171-w |
| [9] |
M. C. Deo, S. S. Jagdale, Prediction of breaking waves with neural networks, Ocean Eng., 30 (2003), 1163–1178. https://doi.org/10.1016/S0029-8018(02)00086-0 doi: 10.1016/S0029-8018(02)00086-0
|
| [10] | D. Yu, X. Li, X. Wang, W. Huang, X. Hu, Y. Jia, Community modularity structure promotes the evolution of phase clusters and chimeralike states, Phys. Rev. E, 111 (2025). https://doi.org/10.1103/PhysRevE.111.034311 |
| [11] |
Y. Xu, J. Ma, Pattern formation in a thermosensitive neural network, Commun. Nonlinear Sci. Numer. Simul., 111 (2022), 106426. https://doi.org/10.1016/j.cnsns.2022.106426 doi: 10.1016/j.cnsns.2022.106426
|
| [12] |
R. Wang, J. Li, M. Du, J. Lei, Y. Wu, Transition of spatiotemporal patterns in neuronal networks with chemical synapses, Commun. Nonlinear Sci. Numer. Simul. 40 (2016), 80–88. https://doi.org/10.1016/j.cnsns.2016.04.018 doi: 10.1016/j.cnsns.2016.04.018
|
| [13] |
J. Ma, J. Tang, A review for dynamics of collective behaviors of network of neurons, Sci. China Inf. Sci., 58 (2015), 2038–2045. https://doi.org/10.1007/s11431-015-5961-6 doi: 10.1007/s11431-015-5961-6
|
| [14] |
X. Liu, Y. Yu, Q. Wang, Dynamic epileptic seizure propagation based on multiscale synaptic plasticity, Nonlinear Dyn., 113 (2025), 10445–10459. https://doi.org/10.1007/s11071-024-10590-2 doi: 10.1007/s11071-024-10590-2
|
| [15] |
Y. Yu, H. Wang, X. Liu, Q. Wang, Closed-loop transcranial electrical stimulation for inhibiting epileptic activity propagation: A whole-brain model study, Nonlinear Dyn., 112 (2024), 21369–21387. https://doi.org/10.1007/s11071-024-10132-w doi: 10.1007/s11071-024-10132-w
|
| [16] |
A. E. Pereda, Electrical synapses and their functional interactions with chemical synapses, Nat. Rev. Neurosci., 15 (2014), 250–263. https://doi.org/10.1038/nrn3708 doi: 10.1038/nrn3708
|
| [17] |
N. Kopell, B. Ermentrout, Chemical and electrical synapses perform complementary roles in the synchronization of interneuronal networks, Proc. Natl. Acad. Sci. U. S. A., 101 (2004), 15482–15487. https://doi.org/10.1073/pnas.040634310 doi: 10.1073/pnas.040634310
|
| [18] |
P. Ge, H. Cao, Synchronization of Rulkov neuron networks coupled by excitatory and inhibitory chemical synapses, Chaos, 29 (2019), 023129. https://doi.org/10.1063/1.5053908 doi: 10.1063/1.5053908
|
| [19] |
R. Harris-Warrick, Synaptic chemistry in single neurons: GABA is identified as an inhibitory neurotransmitter, J. Neurophysiol., 93 (2005), 3029–3031. https://doi.org/10.1152/classicessays.00026.2005 doi: 10.1152/classicessays.00026.2005
|
| [20] |
D. Guo, Q. Wang, M. Perc, Complex synchronous behavior in interneuronal networks with delayed inhibitory and fast electrical synapses, Phys. Rev. E, 85 (2012), 061905. https://doi.org/10.1103/PhysRevE.85.061905 doi: 10.1103/PhysRevE.85.061905
|
| [21] |
Y. Shao, F. Wu, Q. Wang, Synchronization and complex dynamics in locally active threshold memristive neurons with chemical synapses, Nonlinear Dyn., 112 (2024), 13483–13502. https://doi.org/10.1007/s11071-024-09747-w doi: 10.1007/s11071-024-09747-w
|
| [22] |
Y. Jia, H. Gu, X. Wang, Nonlinear mechanisms for enhanced and synchronized post-inhibitory rebound spiking associated with seizures in an inhibitory–excitatory neuronal network, Chaos, 35 (2025), 033104. https://doi.org/10.1063/5.0232718 doi: 10.1063/5.0232718
|
| [23] |
R. Wang, H. Gu, Y. Li, Nonlinear mechanism for paradoxical facilitation of spike induced by inhibitory synapse in auditory nervous system for sound localization, Nonlinear Dyn., 112 (2024), 19393–19419. https://doi.org/10.1007/s11071-024-10008-z doi: 10.1007/s11071-024-10008-z
|
| [24] |
P. Zhou, Y. Xu, J. Ma, Dynamical and coherence resonance in a photoelectric neuron under autaptic regulation, Physica A, 620 (2023), 128746. https://doi.org/10.1016/j.physa.2023.128746 doi: 10.1016/j.physa.2023.128746
|
| [25] |
X. Li, Y. Xu, How lights affect the circadian rhythm in sleep-awake circle, Chin. J. Phys., 91 (2024), 719–733. https://doi.org/10.1016/j.cjph.2024.08.016 doi: 10.1016/j.cjph.2024.08.016
|
| [26] |
X. Li, Y. Xu, Energy level transition and mode transition in a neuron, Nonlinear Dyn., 112 (2024), 2253–2263. https://doi.org/10.1007/s11071-023-09147-6 doi: 10.1007/s11071-023-09147-6
|
| [27] |
F. Yang, J. Ma, G. Ren, A Josephson junction-coupled neuron with double capacitive membranes, J. Theor. Biol., 578 (2024), 111686. https://doi.org/10.1016/j.jtbi.2023.111686 doi: 10.1016/j.jtbi.2023.111686
|
| [28] |
Y. Guo, Y. Xie, J. Ma, Nonlinear responses in a neural network under spatial electromagnetic radiation, Physica A, 626 (2023), 129120. https://doi.org/10.1016/j.physa.2023.129120 doi: 10.1016/j.physa.2023.129120
|
| [29] |
Y. Wu, Q. Ding, W. Huang, X. Hu, Z. Ye, Y. Jia, Dynamic modulation of external excitation enhance synchronization in complex neuronal network, Chaos Soliton. Fract., 183 (2024), 114896. https://doi.org/10.1016/j.chaos.2024.114896 doi: 10.1016/j.chaos.2024.114896
|
| [30] | W. Huang, Y. Wu, Q. Ding, Y. Jia, Effects of potassium channel blockage on chimera-like states in the excitatory–inhibitory neuronal network, Eur. Phys. J. Spec. Top., (2025)https://doi.org/10.1140/epjs/s11734-025-01529-8 |
| [31] |
T. Obut, E. Cimen, M. Cakir, A novel numerical approach for solving delay differential equations arising in population dynamics, Math. Modell. Control, 3 (2023), 233–243. https://doi.org/10.3934/mmc.2023020 doi: 10.3934/mmc.2023020
|
| [32] |
Y. Jia, H. Gu, Y. Li, Influence of inhibitory autapses on synchronization of inhibitory network gamma oscillations, Cognitive Neurodyn., 17 (2023), 1131–1152. https://doi.org/10.1007/s11571-022-09856-5 doi: 10.1007/s11571-022-09856-5
|
| [33] |
Q. Wang, M. Perc, Z. Duan, G. Chen, Delay-induced multiple stochastic resonances on scale-free neuronal networks, Chaos, 19 (2009), 325407. https://doi.org/10.1063/1.3133126 doi: 10.1063/1.3133126
|
| [34] |
K. Xu, J. P. Maidana, P. Orio, Diversity of neuronal activity is provided by hybrid synapses, Nonlinear Dyn., 105 (2021), 2693–2710. https://doi.org/10.1007/s11071-021-06704-9 doi: 10.1007/s11071-021-06704-9
|
| [35] |
J. T. Fossi, Z. T. Njitacke, W. N. Tankeu, J. M. Mendimi, J. Awrejcewicz, J. Atangana, Phase synchronization and coexisting attractors in a model of three different neurons coupled via hybrid synapses, Chaos Soliton. Fract., 177 (2023), 114202. https://doi.org/10.1016/j.chaos.2023.114202 doi: 10.1016/j.chaos.2023.114202
|
| [36] |
D. Yu, X. Zhou, G. Wang, Q. Ding, T. Li, Y. Jia, Effects of chaotic activity and time delay on signal transmission in FitzHugh-Nagumo neuronal system, Cognitive Neurodyn., 16 (2022), 887–897. https://doi.org/10.1007/s11571-021-09743-5 doi: 10.1007/s11571-021-09743-5
|
| [37] |
F. Yang, J. Ma, F. Wu, Review on memristor application in neural circuit and network, Chaos Soliton. Fract., 187 (2024), 115361. https://doi.org/10.1016/j.chaos.2024.115361 doi: 10.1016/j.chaos.2024.115361
|
| [38] |
M. Sainz-Trapaga, C. Masoller, H. A. Braun, M. T. Huber, Influence of time-delayed feedback in the firing pattern of thermally sensitive neurons, Physical Review E, 70 (2004), 031904. https://doi.org/10.1103/PHYSREVE.70.031904 doi: 10.1103/PHYSREVE.70.031904
|
| [39] |
X. J. Wang, G. Buzsáki, Gamma oscillation by synaptic inhibition in a hippocampal interneuronal network model, J. Neurosci., 16 (1996), 6402–6413. https://doi.org/10.1186/1471-2202-12-S1-P264 doi: 10.1186/1471-2202-12-S1-P264
|
| [40] |
A. L. Hodgkin, A. F. Huxley, Propagation of electrical signals along giant nerve fibres, Proc. R. Soc. B, 140 (1952), 177–183. https://doi.org/10.1098/rspb.1952.0054 doi: 10.1098/rspb.1952.0054
|
| [41] |
A. L. Hodgkin, The Croonian Lecture-Ionic movements and electrical activity in giant nerve fibres. Proceedings of the Royal Society of London, Proc. R. Soc. B, 148 (1958), 1–37. https://doi.org/10.1098/rspb.1958.0001 doi: 10.1098/rspb.1958.0001
|
| [42] |
R. D. Traub, N. Kopell, A. Bibbig, E. H. Buhl, F. E. LeBeau, M. A. Whittington, Gap junctions between interneuron dendrites can enhance synchrony of gamma oscillations in distributed networks, J. Neurosci., 21 (2001), 9478–9486. https://doi.org/10.1523/JNEUROSCI.21-23-09478.2001 doi: 10.1523/JNEUROSCI.21-23-09478.2001
|
| [43] |
M. Bartos, I. Vida, M. Frotscher, A. Meyer, H. Monyer, J. R. Geiger, et al., Fast synaptic inhibition promotes synchronized gamma oscillations in hippocampal interneuron networks, Proc. Natl. Acad. Sci. U. S. A., 99 (2002), 13222–13227. https://doi.org/10.1073/pnas.192233099 doi: 10.1073/pnas.192233099
|
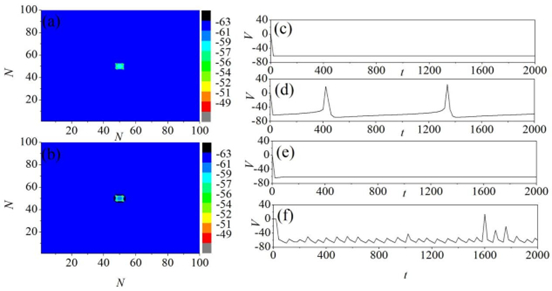
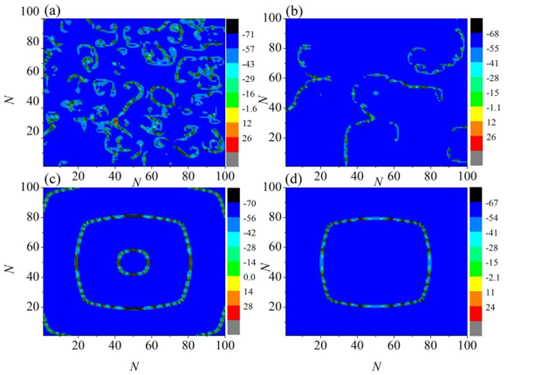
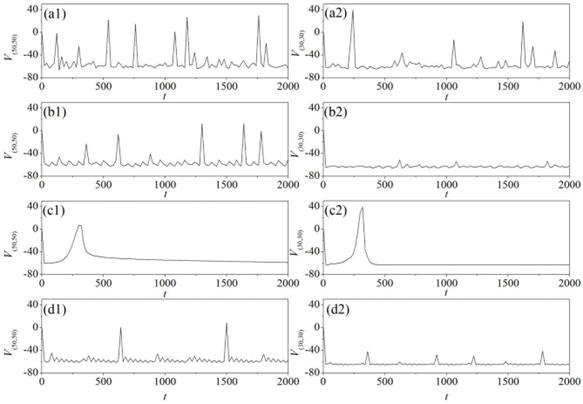
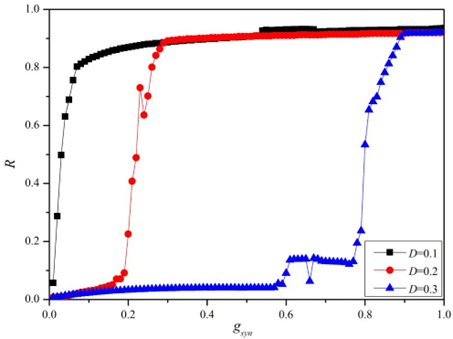
Figures(11)

Ying Xu, Xiaodi Li. The pattern dynamics of interneuronal networks with inhibitory synaptic coupling[J]. AIMS Mathematics, 2025, 10(5): 10976-10993. doi: 10.3934/math.2025498







 DownLoad:
DownLoad: