Citation: Halis Bilgil. New grey forecasting model with its application and computer code[J]. AIMS Mathematics, 2021, 6(2): 1497-1514. doi: 10.3934/math.2021091

| [1] |
J. L. Deng, Control problems of grey systems, Syst. Control Lett., 1 (1982), 288-294. doi: 10.1016/S0167-6911(82)80025-X

|
| [2] |
L. F. Wu, S. F. Liu, D. Chen, L. Yao, W. Cui, Using gray model with fractional order accumulation to predict gas emission, Nat. Hazards, 71 (2014), 2231-2236. doi: 10.1007/s11069-013-0960-z
|
| [3] | Y. G. Zhang, Y. Xu, Z. P. Wang, GM (1, 1) grey prediction of lorenz chaotic system, Chaos Solitons Fract., 42 (2009), 1003-1009. |
| [4] | W. Zhou, J. M. He, Generalized GM (1, 1) model and its application in forecasting of fuel production, Appl. Math. Modell., 37 (2013), 6234-6243. |
| [5] | S. A. Javed, S. Liu, Predicting the research output/growth of selected countries: application of even GM (1, 1) and ndgm models, Scientometrics, 115 (2018), 395-413. |
| [6] |
S. L. Ou, Forecasting agricultural output with an improved grey forecasting model based on the genetic algorithml, Comput. Electron. Agric., 85 (2012), 33-39. doi: 10.1016/j.compag.2012.03.007
|
| [7] | B. Zeng, Y. Tan, H. Xu, J. Quan, L. Wang, X. Zhou, Forecasting the electricity consumption of commercial sector in hong kong using a novel grey dynamic prediction model, J. Grey Syst., 30 (2018), 157-172. |
| [8] | S. Ene, N. Ozturk, Grey modelling based forecasting system for return flow of end-of-life vehicles, Technol. Forecast. Soc. Change, 117 (2018), 155-166. |
| [9] | U. Şahin, T. Şahin, Forecasting the cumulative number of confirmed cases of COVID-19 in italy, uk and usa using fractional nonlinear grey bernoulli model, Chaos Solitons Fract., 138 (2020), 109948. |
| [10] |
X. Ma, Z. B. Liu, Application of a novel time-delayed polynomial grey model to predict the natural gas consumption in china, J. Comput. Appl. Math., 324 (2017), 17-24. doi: 10.1016/j.cam.2017.04.020
|
| [11] |
J. Cui, S. Liu, B. Zeng, N. Xie, A novel grey forecasting model and its optimization, Appl. Math. Model., 37 (2013), 4399-4406. doi: 10.1016/j.apm.2012.09.052
|
| [12] |
X. Ma, Z. Liu, Y. Wang, Application of a novel nonlinear multivariate grey bernoulli model to predict the tourist income of china, J. Comput. Appl. Math., 347 (2019), 84-94. doi: 10.1016/j.cam.2018.07.044
|
| [13] | Z. X. Wang, Q. Li, L. L. Pei, A seasonal GM (1, 1) model for forecasting the electricity consumption of the primary economic sectors, Energy, 154 (2018), 522-534. |
| [14] |
L. Z. Wu, S. H. Li, R. Q. Huang, Q. Xi, A new grey prediction model and its application to predicting landslide displacement, Appl. Soft Comput., 95 (2020), 106543. doi: 10.1016/j.asoc.2020.106543
|
| [15] |
L. Liu, L. Wu, Forecasting the renewable energy consumption of the european countries by an adjacent non-homogeneous grey model, Appl. Math. Modell., 89 (2021), 1932-1948. doi: 10.1016/j.apm.2020.08.080
|
| [16] |
L. Ye, N. Xie, A. Hu, A novel time-delay multivariate grey model for impact analysis of CO2 emissions from chinas transportation sectors, Appl. Math. Modell., 91 (2021), 493-507. doi: 10.1016/j.apm.2020.09.045
|
| [17] |
W. Wu, X. Ma, Y. Wang, W. Cai, B. Zeng, Predicting chinas energy consumption using a novel grey riccati model, Appl. Soft Comput., 95 (2020), 106555. doi: 10.1016/j.asoc.2020.106555
|
| [18] |
K. D. Yin, Y. Geng, X. M. Li, Improved grey prediction model based on exponential grey action quantity, J. Syst. Eng. Electron., 29 (2018), 560-570. doi: 10.21629/JSEE.2018.03.13
|
| [19] |
D. Akay, M. Atak, Grey prediction with rolling mechanism for electricity demand forecasting of Turkey, Energy, 32 (2007), 1670-1675. doi: 10.1016/j.energy.2006.11.014
|
| [20] |
J. Y. Chiang, C. K. Chen, Application of grey prediction to inverse nonlinear heat conduction problem, Int. J. Heat Mass Transfer, 51 (2008), 576-585. doi: 10.1016/j.ijheatmasstransfer.2007.05.015
|
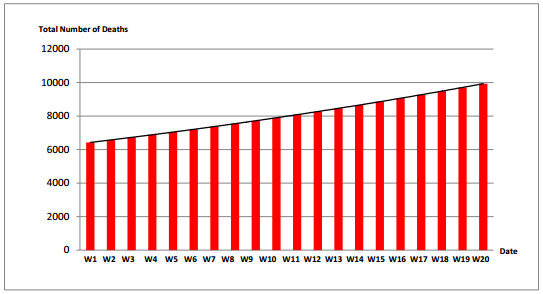
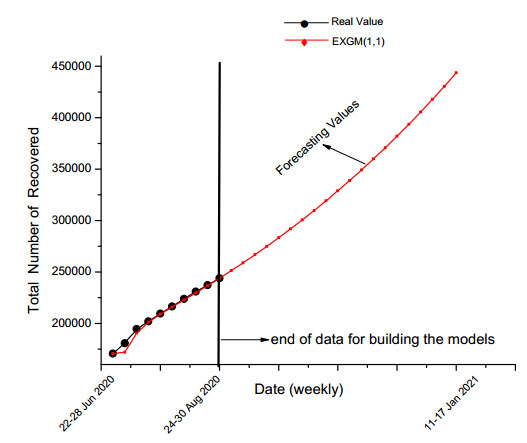
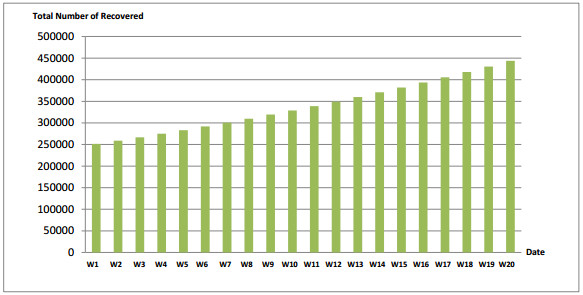
Figures(7) / Tables(7)

Halis Bilgil. New grey forecasting model with its application and computer code[J]. AIMS Mathematics, 2021, 6(2): 1497-1514. doi: 10.3934/math.2021091







 DownLoad:
DownLoad: