Citation: Xiangjun Dai, Suli Wang, Baoping Yan, Zhi Mao, Weizhi Xiong. Survival analysis of single-species population diffusion models with chemotaxis in polluted environment[J]. AIMS Mathematics, 2020, 5(6): 6749-6765. doi: 10.3934/math.2020434

| [1] | E. H. Colombo, C. Anteneodo, Metapopulation dynamics in a complex ecological landscape, Phys. Rev. E, 92 (2015), 022714. |
| [2] | E. Beretta, F. Solimano, Y. Takeuchi, Global stability and periodic orbits for two-patch predatorprey diffusion-delay models, Math. Biosci., 85 (1987), 153-183. |
| [3] | G. Z. Zeng, L. S. Chen, J. F. Chen, Persistence and periodic orbits for two-species nonautonomous diffusion lotka-volterra models, Math. Comput. Model., 20 (1994), 69-80. |
| [4] | E. Beretta, Y. Takeuchi, Global stability of single-species diffusion Volterra models with continuous time delays, B. Math. Biol., 49 (1987), 431-448. |
| [5] | L. Zhang, Z. Teng, The dynamical behavior of a predator-prey system with Gompertz growth function and impulsive dispersal of prey between two patches, Math. Method. Appl. Sci., 39 (2016), 3623-3639. |
| [6] | L. J. S. Allen, Persistence and extinction in single-species reaction-diffusion models, B. Math. Biol., 45 (1983), 209-227. |
| [7] | H. I. Freedman, Single species migration in two habitats: Persistence and extinction, Math. Model., 8 (1987), 778-780. |
| [8] | M. Bengfort, H. Malchow, F. M. Hilker, The Fokker-Planck law of diffusion and pattern formation in heterogeneous environments, J. Math. Biol., 73 (2016), 683-704. |
| [9] | D. Li, S. J. Guo, Stability and Hopf bifurcation in a reaction-diffusion model with chemotaxis and nonlocal delay effect, Int. J. Bifurcat. Chaos, 28 (2018), 1850046. |
| [10] | Y. Tan, C. Huang, B. Sun, et al. Dynamics of a class of delayed reaction-diffusion systems with Neumann boundary condition, J. Math. Anal. Appl., 458 (2018), 1115-1130. |
| [11] | Y. Xie, Q. Li, K. Zhu, Attractors for nonclassical diffusion equations with arbitrary polynomial growth nonlinearity, Nonlinear Anal. Real, 31 (2016), 23-37. |
| [12] | F. Wei, S. A. H. Geritz, J. Cai, A stochastic single-species population model with partial pollution tolerance in a polluted environment, Appl. Math. Lett., 63 (2016), 130-136. |
| [13] | F. Y. Wei, L. H. Chen, Psychological effect on single-species population models in a polluted environment, Math. Biosci., 290 (2017), 22-30. |
| [14] | J. J. Jiao, L. S. Chen, The extinction threshold on a single population model with pulse input of environmental toxin in a polluted environment, Math. Appl., 22 (2009), 11-19. |
| [15] | Y. Xiao, L. Chen, Effects of toxicants on a stage-structured population growth model, Appl. Math. Comput., 123 (2001), 63-73. |
| [16] | J. D. Stark, J. E. Banks, Population-level effects of pesticides and other toxicants on arthropods, Annu. Rev. Entomol., 48 (2003), 505-519. |
| [17] | W. Jing, W. Ke, Analysis of a single species with diffusion in a polluted environment, Electron. J. Differ. Eq., 112 (2006), 285-296. |
| [18] | R. M. May, Stability and Complexity in Model Ecosystems, Princeton University Press, 2001. |
| [19] | A. Friedman, Stochastic Differential Equations and Applications, Academic Press, 1976. |
| [20] | M. Liu, K. Wang, Persistence and extinction in stochastic non-autonomous logistic systems, J. Math. Anal. Appl., 375 (2011), 443-457. |
| [21] | X. J. Dai, Z. Mao, X. J. Li, A stochastic prey-predator model with time-dependent delays, Adv. Differ. Equ., 2017 (2017), 1-15. |
| [22] | M. Liu, K. Wang, Q. Wu, Survival analysis of stochastic competitive models in a polluted environment and stochastic competitive exclusion principle, B. Math. Biol., 73 (2011), 1969-2012. |
| [23] | X. Zou, D. Fan, K. Wang, Effects of dispersal for a logistic growth population in random environments, Abstr. Appl. Anal., 2013 (2013), 1-9. |
| [24] | L. Zu, D. Jiang, ORegan. Donal, Stochastic permanence, stationary distribution and extinction of a single-species nonlinear diffusion system with Rrandom perturbation, Abstr. Appl. Anal., 2014 (2014), 1-14. |
| [25] | M. Liu, M. Deng, B. Du, Analysis of a stochastic logistic model with diffusion, Appl. Comput. Model., 266 (2015), 169-182. |
| [26] | X. Zou, K. Wang, A robustness analysis of biological population models with protection zone, Appl. Math. Model., 35 (2011), 5553-5563. |
| [27] | X. Zou, K. Wang, M. Liu, Can protection zone potentially strengthen protective effects in random environments?, Appl. Math. Comput., 231 (2014), 26-38. |
| [28] | F. Y. Wei, C. J. Wang, Survival analysis of a single-species population model with fluctuations and migrations between patches, Appl. Math. Model., 81 (2020), 113-127. |
| [29] | X. Zou, K. Wang, Dynamical properties of a biological population with a protected area under ecological uncertainty, Appl. Math. Model., 39 (2015), 6273-6284. |
| [30] | D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525-546. |

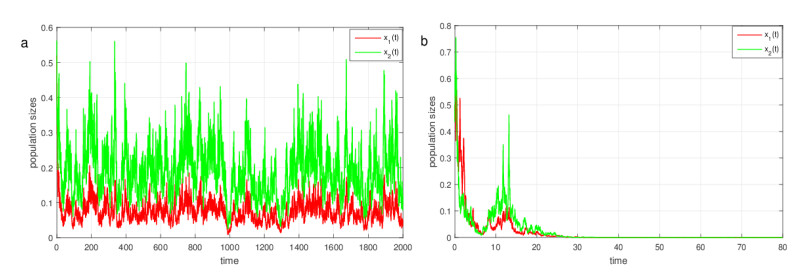
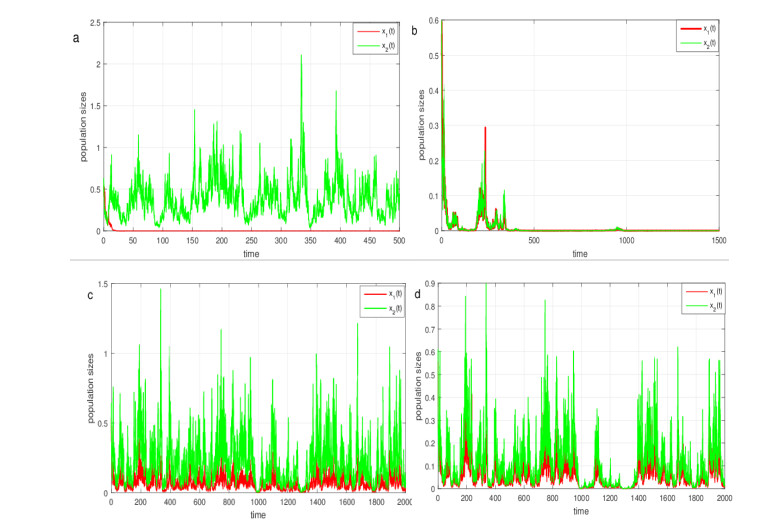
Figures(3)

Xiangjun Dai, Suli Wang, Baoping Yan, Zhi Mao, Weizhi Xiong. Survival analysis of single-species population diffusion models with chemotaxis in polluted environment[J]. AIMS Mathematics, 2020, 5(6): 6749-6765. doi: 10.3934/math.2020434







 DownLoad:
DownLoad: