Citation: Charles Makoundi, Khin Zaw, R.R. Large. Laser Ablation ICPMS Analysis of Pyrite and U-Pb Zircon Dating of Host Rocks From the Tersang Gold Deposit, Malaysia[J]. AIMS Geosciences, 2017, 3(3): 396-437. doi: 10.3934/geosci.2017.3.396

| [1] | Richardson JA (1939) The Geology and mineral resources of the neighbourhood of Raub, Pahang, Federated Malay States, with an account of the Geology of the Raub Australian Gold Mine. Geological Survey Department, Federated Malay States, Memoir 3: 166. |
| [2] |
Makoundi C, Khin Zaw, Large RR, et al. (2014) Geology, geochemistry and metallogenesis of the Selinsing gold deposit, central Malaysia. Gondwana Res 26: 241-261. doi: 10.1016/j.gr.2013.08.023

|
| [3] |
Metcalfe I (2011) Tectonic framework and Phanerozoic evolution of Sundaland. Gondwana Res 19: 3-21. doi: 10.1016/j.gr.2010.02.016
|
| [4] | Metcalfe I (2012) Cold Gondwana to warm Tethys: Late Paleozoic-Mesozoic evolution of Tibetan and SE Asian continental blocks. 2012 IAGR Annual Convention and 9th International Symposium on Gondwana to Asia, Adelaide, Australia. |
| [5] |
Oliver G, Khin Zaw, Hotson Mark, et al. (2014) U-Pb zircon geochronology of Early Permian to Late Triassic rocks from Singapore and Johor: a plate tectonic reinterpretation. Gondwana Res 26: 132-143. doi: 10.1016/j.gr.2013.03.019
|
| [6] |
Khin Zaw, Meffre S, Lai C-K, et al. (2014) Tectonics and metallogeny of mainland southeast Asia - a review and contribution. Gondwana Res 26: 1-4. doi: 10.1016/j.gr.2014.01.005
|
| [7] |
Yeap EB (1993) Tin and gold mineralisation in peninsular Malaysia and their relationships to the tectonic development. J Southeast Asian Earth Sci 8: 329-348. doi: 10.1016/0743-9547(93)90035-N
|
| [8] | Scrivenor JB (1928) The geology of Malayan ore deposits. Macmillan, London, 216. |
| [9] |
Metcalfe I (2013) Tectonic evolution of the Malay Peninsula. J Asian Earth Sci 76: 195-213. doi: 10.1016/j.jseaes.2012.12.011
|
| [10] | Kobayashi T, Tamura M (1968) Myophoria in Malaya with a note on the Triassic Trigoniacea. In: Kobayashi, T and Toriyama, R (Eds), Geology and palaeontology of Southeast Asia, University of Tokyo Press, 5: 88-137. |
| [11] | Makoundi C (2004) Facies analysis of the Triassic Jelai Formation in the Central Basin of Peninsular Malaysia: Implications on Paleogeography and Tectonics. MSc thesis, University of Malaya, 140. |
| [12] | Gobbett DJ, Hutchison CS (Eds) (1973) Geology of the Malay Peninsula: West Malaysia and Singapore. New York: Wiley-Interscience, 438. |
| [13] | Tan BK (1984) The Tectonic framework and evaluation of the Central Belt and its margin, Peninsular Malaysia. Geological Society of Malaysia. Bulletin 17: 307-322. |
| [14] |
Metcalfe I (2002) Permian Tectonic Framework and Paleogeography of SE Asia. J Asian Earth Sci 20: 551-566. doi: 10.1016/S1367-9120(02)00022-6
|
| [15] |
Schwartz MO, Rajah SS, Askury AK, et al. (1995) The southeast Asian tin belt. Earth Sci Rev 38: 95-293 doi: 10.1016/0012-8252(95)00004-T
|
| [16] | Bignell JD, Snelling NJ (1977) K-Ar ages on some basic igneous rocks from peninsular Malaysia and Thailand. Bull Geol Soc Malays 8: 89-93. |
| [17] |
Metcalfe I (2000) The Bentong-Raub Suture Zone. J Asian Earth Sci 18: 691-712. doi: 10.1016/S1367-9120(00)00043-2
|
| [18] | Kosler J (2001) Laser-ablation ICPMS study of metamorphic minerals and processes. Mineral Assoc Can Short Course Handb 29: 185-202. |
| [19] | Large RR, Danyushevsky L, Hollit C, et al. (2009) Gold and trace element zonation in pyrite using a laser imaging technique: implications for the timing of gold in orogenic and Carlin-style sediment-hosted deposits: Econ Geol 104: 635-668. |
| [20] |
Danyushevsky LV, Robinson P, Gilbert S, et al. (2011) Routine quantitative multi-element analysis of sulphide minerals by laser ablation ICP-MS: Standard development and consideration of matrix effects. Geochemistry: Exploration, Environment, And Analysis 11: 51-60. doi: 10.1144/1467-7873/09-244
|
| [21] | Wiendenbeck M, Alle P, Corfu F, et al. (1995) Three Natural zircon standards for U-Th-Pb, Lu-Hf, trace-element and REE analyses. Geostand Newsl 19: 1-23. |
| [22] | Black LP, Kamos L, Allen CM, et al. (2003) TEMORA 1. A new zircon standard for Phanerozoic U-Pb geochronology. Chem Geol 200: 155-170. |
| [23] |
Jackson SE, Pearson NJ, Griffin WL, et al. (2004) The application of laser ablation inductively coupled plasma-mass spectrometry to in situ U-Pb zircon geochronology. Chem Geol 211: 47-69. doi: 10.1016/j.chemgeo.2004.06.017
|
| [24] | Black LP, Gulson BL (1978) The age of the Mud Tank Carbonatite, Strangways Range, Northern Territory. BMR J Aust Geol Geophys 3: 227-232. |
| [25] |
Black LP, Kamo SL, Allen CM, et al. (2004) Improved 206Pb/238U microprobe geochronology by the monitoring of a trace-element related matrix effect; SHRIMP, ID-TIMS, ELA-ICP-MS, and oxygen isotope documentation for a series of zircon standards. Chem Geol 205: 115-140. doi: 10.1016/j.chemgeo.2004.01.003
|
| [26] |
Meffre S, Large RR, Scott R, et al. (2008) Age and pyrite Pb-isotopic composition of the giant Sukhoi Log sediment-hosted gold deposit, Russia. Geochimt Cosmochim Acta 72: 2377-2391. doi: 10.1016/j.gca.2008.03.005
|
| [27] | Paton C, Woodhead JD, Hellstrom JC, et al. (2010) Improved laser ablation U-Pb zircon geochronology through robust down-hole fractionation correction. Geochem Geophys Geosyst 11: 1525-2027. |
| [28] | Rumsey DJ (2016) Statistics For Dummies. 2nd Edition, Wiley Brand, 48. |
| [29] | Haile NS, Stauffer PH, Krishnan D, et al. (1977) Paleozoic redbeds and radiolarian chert: reinterpretation of their relationships in the Bentong and Raub areas, West Pahang, peninsular Malaysia. Geol Soc Malays Bull 8: 45-60. |
| [30] |
Hoskin PWO, Schaltegger U (2003) The composition of zircon and igneous and metamorphic petrogenesis. Rev mineral geochem 53: 27-62. doi: 10.2113/0530027
|
| [31] |
Chun-Kit Lai, Meffre S, Crawford AJ, et al. (2014) The Central Ailaoshan ophiolite and modern analogues. Gondwana Res 26: 75-88. doi: 10.1016/j.gr.2013.03.004
|
| [32] |
Large RR, Bull SW, Maslennikov VV (2011) A carbonaceous sedimentary source-rock model for Carlin-type and orogenic gold deposits. Soc Econ Geol 106: 331-358. doi: 10.2113/econgeo.106.3.331
|
| [33] | Kolker A, Cannon WF, Westjohn DB, et al. (1998) Arsenic-rich pyrite in the Mississippian Marshall Sandstone: source of anomalous arsenic in southeatern Michigan ground water. Geol Soc Am Abstr Progr 30: A-59. |
| [34] | Kolker A, Goldhaber MB, Hatch JR, et al. (1999) Arsenic-rich pyrite in coals of the Warrior Field, northwestern Alabama: mineralogical evidence for a hydrothermal origin. Geol Am Abstr Progr 31: A-402. |
| [35] |
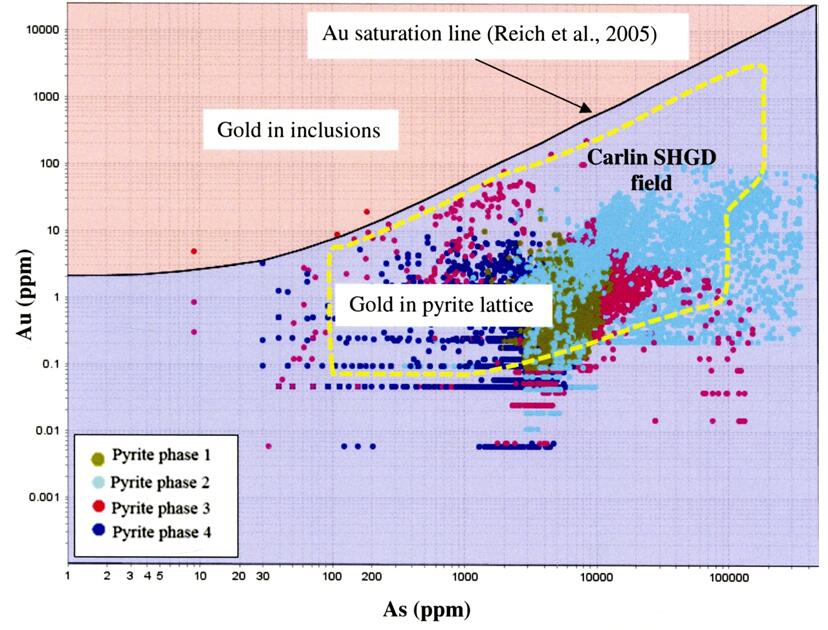
Reich M, Kesler SE, Utsunomiya S, et al. (2005) Solubility of gold in arsenian pyrite. Geochem Cosmochim Acta 69: 2781-2796. doi: 10.1016/j.gca.2005.01.011
|
| [36] | Goh KC (2013) Geological setting and mineralisation characteristics of the sedimentary rock hosted disseminated Jugan Au deposit, Bau mining district, Sarawak, Malaysia. BSc (Honours) thesis, University of Tasmania, 94. |



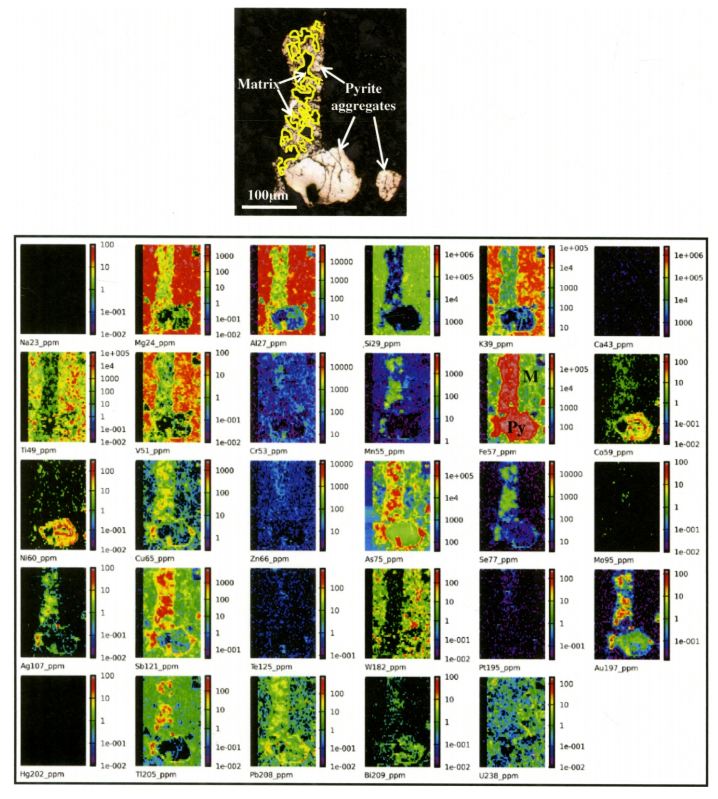
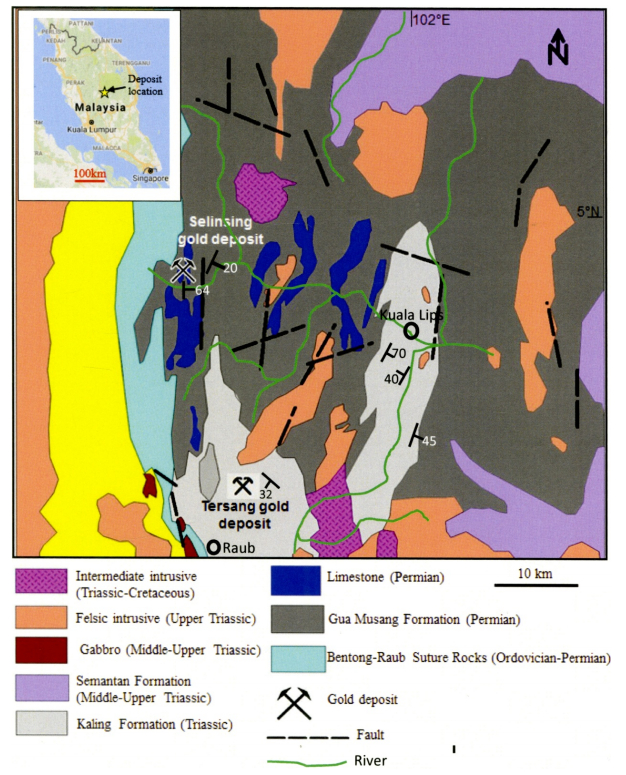
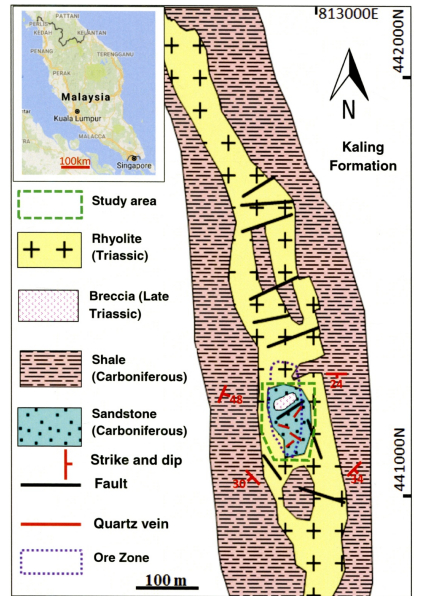


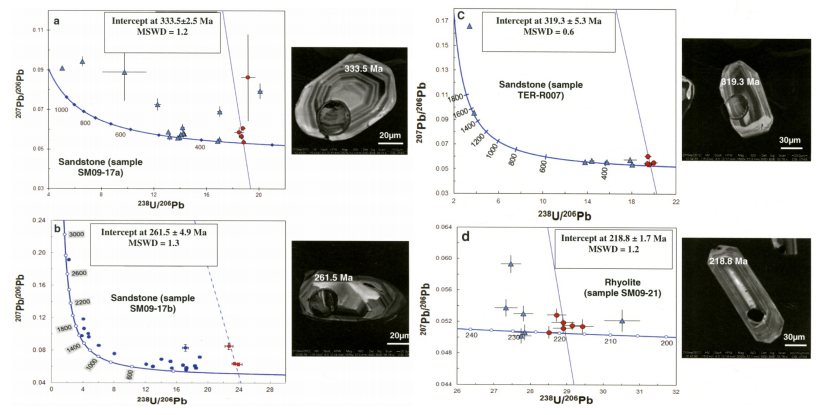


Figures(20) / Tables(12)

Charles Makoundi, Khin Zaw, R.R. Large. Laser Ablation ICPMS Analysis of Pyrite and U-Pb Zircon Dating of Host Rocks From the Tersang Gold Deposit, Malaysia[J]. AIMS Geosciences, 2017, 3(3): 396-437. doi: 10.3934/geosci.2017.3.396







 DownLoad:
DownLoad: