Citation: John G. Rogers, Samuel J.G. Cooper, Simone Cooper, Danielle Densley Tingley, Naomi Braithwaite, Mariale Moreno, Alex Rodrigues, and Giuseppe Salvia. Product longevity and shared ownership: Sustainable routes to satisfying the world’s growing demand for goods [J]. AIMS Energy, 2015, 3(4): 547-561. doi: 10.3934/energy.2015.4.547

| [1] | Kharas H (2010) The Emerging Middle Class In Developing Countries, OECD Global Development Outlook, Working Paper No. 285. |
| [2] | UN, Report of the Conference of the Parties on its eighteenth session, held in Doha from 26 November to 8 December 2012, FCCC/CP/2012/8, 2012. Available from: http://unfccc.int/resource/docs/2012/cop18/eng/08.pdf Aug 2013 |
| [3] | International Energy Agency, Energy Technology Transitions for Industry—Strategies for the next industrial revolution , IEC/OECD 2009 |
| [4] |
Dyer CH, Hammond GP, Jones CI, et al. (2008) Enabling technologies for industrial energy demand management. Energy Policy 36: 4434-4443. doi: 10.1016/j.enpol.2008.09.028

|
| [5] |
Allwood JM, Ashby MF, Gutowski TG, et al. (2013) Material efficiency: providing material services with less material production, rsta.royalsocietypublishing.org Phil Trans R Soc A 371: 20120496. doi: 10.1098/rsta.2012.0496
|
| [6] | McKinsey & Company (2012) Resource Revolution: Meeting the worlds energy, material, food and water need. Available from: http:/www.mckinsey.com/insights/energy_resource_material/resource_revolution. |
| [7] | Allwood JM, Cullen JM, Carruth MA, et al. (2014) Sustainable Materials with both Eyes Open, UTI Cambridge England. |
| [8] | International Energy Agency, World Energy Outlook 2013. Available from: www.worldenergyoutlook.org |
| [9] | Leibling D (2008) Car ownership in Great Britain, Royal Automobile Club Foundation for Motoring. Available from: http://www.racfoundation.org/assets/rac_foundation/content/downloadables/car%20ownership%20in%20great%20britain%20-%20leibling%20-%20171008%20-%20report.pdf (accessed Sept 2014). |
| [10] |
Nieuwenhuis P (2008) From banger to classic—a model for sustainable car consumption? Int J Consum Stud 32: 648-655. doi: 10.1111/j.1470-6431.2008.00721.x
|
| [11] | Nieuwenhuis P (1994) The long life car: investigation a motor industry heresy, in Motor vehicles in the environment: principle and practice ed Nieuwenhuis P & Wells P, John Wiley and sons, Chichester. |
| [12] | Smart Moves Ltd, London City Car Club Surveys Summary of Key Points, Report for The London City Car Club Consortium, 2004. |
| [13] |
Cooper T (2005) Slower consumption; reflections on product life spans and the ‘Throwaway Society'. J Ind Ecol 9: 51-67. doi: 10.1162/1088198054821609
|
| [14] |
Goodall P, Rosamond E, Harding J (2014) A review of the state of the art in tools and techniques used to evaluate remanufacturing feasibility. J Clean Prod 81: 1-15 doi: 10.1016/j.jclepro.2014.06.014
|
| [15] | Ijomah W L,McMahon CA (2007) Hammond G P,Newman S T, Development of design for remanufacturing guidelines to support sustainable manufacturing Robotics and Computer-Integrated Manufacturing 23: 712-719. |
| [16] | Centre for remanufacturing and reuse web site. Available from: http://www.remanufacturing.org.uk/policy-sector.lasso (accessed Sept 2014). |
| [17] | S Allen (2010) Drivers and barriers for remanufacturing of small- and medium-scale wind turbines, Centre for Remanufacturing and Reuse Product Group Study. Available from: http://www.remanufacturing.org.uk/pdf/story/. |
| [18] | Rolls Royce web site. Available from: http://www.rolls-royce.com/sustainability/better_power/products/index.jsp (accessed Sept 2014). |
| [19] | Densley Tingley D, Allwood JM (2014) Reuse of structural steel: the opportunities and challenges, presented at European steel environment & energycongress 2014, Teesside University, UK. |
| [20] | Densley Tingley D Reducing Material Demand in Construction Indemand U A Prospectus, Department of Engineering, University of Cambridge Trumpington Street, Cambridge CB2 1PZ, United Kingdom. Available from: http://www.ukindemand.ac.uk/sector/construction Sept 2014. |
| [21] | Hart GH (2007) A Good Business Move: Recycling Insulation Materials, Insulation Outlook. |
| [22] | Foresight (2013) The Future of Manufacturing: A new era of opportunity and challenge for the UK Summary Report, The Government Office for Science, London. |
| [23] | Deloitte Research (2006) The Service Revolution in Global Manufacturing Industries, A Deloitte Research Global Manufacturing Study. Available from: http://www.apec.org.au/docs/2011-11_training/deloitte2006.pdf (Accessed on Sept 2014). |
| [24] | Visnjic I, Van Looy B (2011) Can a Product Manufacturer Become a Successful Service Provide r? In Pursuit of a Business Model that Fosters Complementarity between Product and Service Activities Perspectives, paper presented at the Academy of Management Conference, San Antonio, USA. Available from: http://www.cambridgeservicealliance.org/uploads/downloadfiles/Can%20a%20Product%20Manufacturer%20be%20a%20Successful%20Service%20Provider.pdf (Accessed on Sept 2014). |
| [25] | Ellen MacArthur Foundation. Towards the circular economy vol.3: Accelerating the scale-up across global supply chains. Available from: http:// www.thecirculareconomy.org (Accessed on July 4, 2014). |
| [26] | Benton D, Hazell J (2014) Wasted opportunities: Smarter systems for resource recovery. A report from the Circular Economy Task Force, Green Alliance, London. Available from: http://www.greenalliance.org.uk/wasted_opportunities:smarter_systems_for_resource_recovery. |
| [27] | European Commission, COMMISSION STAFF WORKING DOCUMENT. ‘Advancing Manufacturin—Advancing Europe’—Report of the Task Force on Advanced Manufacturing for Clean Production, Brussels, 19.3.2014, SWD Final 2014 |
| [28] |
Wells P (2013) Sustainable business models and the automotive industry: A commentary. IIMB Manage Rev 25: 228-239 doi: 10.1016/j.iimb.2013.07.001
|
| [29] |
Williams A (2006) Product-service systems in the automotive industry: the case of micro-factory retailing. J Clean Prod 14: 172-184 doi: 10.1016/j.jclepro.2004.09.003
|
| [30] | UKINDEMAND report on event page web site, Industry seminar on optimizing product lifetimes. Available from: http://www.ukindemand.ac.uk/events/industry-seminar-optimising-product-lifetimes (accessed Sept 2014). |
| [31] | Bjoern R, Nidhi A (2008) Qualitative research skills workshop—a facilitators reference manual, Produced by the Recoup consortium. Available from: htt\\ceid.educ.cam.ac.uk/Publications/RECOUP_Manual.pdf. |
| [32] | ING Economics Department (2015) Rethinking finance in a circular economy. Available from: http://www.ing.com/About-us/Our-stories/Features/Circular-economy-challenges-financial-business-models.htm. |
| [33] | McKinsey Center for Business and Environment (2015) Growth Within: A Circular Economy Vision For A Competitive Europe - a report produced for The Ellen MacArthur Foundation. Available from: http://www.mckinsey.com/~/media/mckinsey/dotcom/client_service/sustainability/2015%20circular%20growth%20report/growth_within-a_circular_economy_vision_for_a_competitive_europe%20(1).ashx. |
| [34] | Cooper T, Braithwaite N, Moreno M, et al. (2015) Product Lifetimes And The Environment Conference Proceedings 17-19 June, 2015 - Nottingham, UK, ISBN 978-0-9576009-9-7 (ebk.)34.Available from: http://www.ntu.ac.uk/plate_conference/PLATE_2015_proceedings.pdf. |
| [35] | OECD (2008) Measuring material flows and resource productivity Volume I. The OECD Guide. Available from: http://www.oecd.org/environment/indicators-modelling-outlooks/MFA-Guide.pdf. |
| [36] | anon (2008) Material Intensity Of The Economy methodology sheet. Available from: http://www.un.org/esa/sustdev/natlinfo/indicators/methodology_sheets/consumption_production/material_intensity.pdf. |
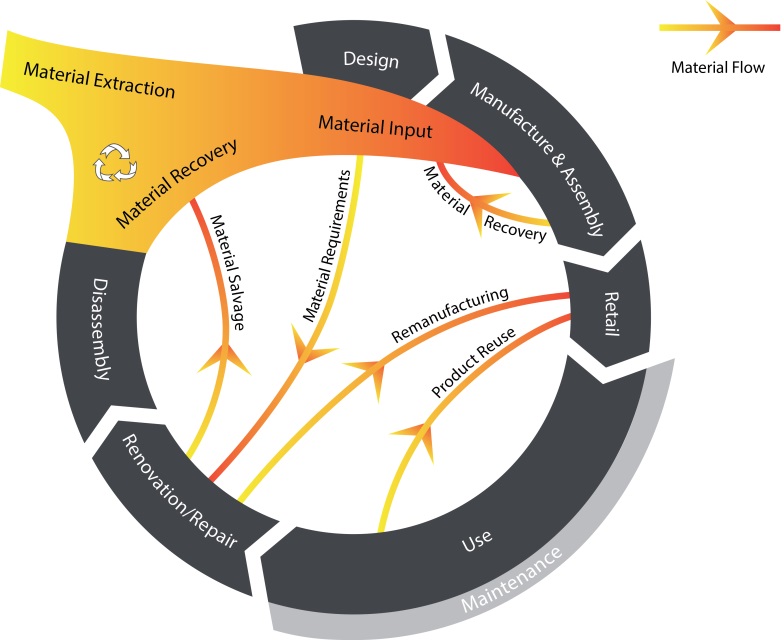
Figures(1) / Tables(3)

John G. Rogers, Samuel J.G. Cooper, Simone Cooper, Danielle Densley Tingley, Naomi Braithwaite, Mariale Moreno, Alex Rodrigues, and Giuseppe Salvia. Product longevity and shared ownership: Sustainable routes to satisfying the world’s growing demand for goods [J]. AIMS Energy, 2015, 3(4): 547-561. doi: 10.3934/energy.2015.4.547







 DownLoad:
DownLoad: