Citation: Michèle Siek, Berenice Marg, Chris M. Ehring, Derya Kirasi, Michael Liebthal, Thorsten Seidel. Interplay of vacuolar transporters for coupling primary and secondary active transport[J]. AIMS Biophysics, 2016, 3(4): 479-500. doi: 10.3934/biophy.2016.4.479

| [1] |
Owens T, Poole RJ (1979) Regulation of cytoplasmic and vacuolar volumes by plant cells in suspension culture. Plant Physiol 64: 900–904. doi: 10.1104/pp.64.5.900

|
| [2] |
Seidel T, Siek M, Marg B, et al. (2013) Energization of vacuolar transport in plant cells and its significance under stress. Intern Rev Cell Mol Biol 304: 57–131. doi: 10.1016/B978-0-12-407696-9.00002-6
|
| [3] |
Tuteja N, Mahajan S (2007) Calcium signaling network in plants. Plant Signal Behav 2: 79–85. doi: 10.4161/psb.2.2.4176
|
| [4] | Hirschi K (2001) Vacuolar H+/Ca2+-transport: who’s directing the traffic? Trends Plant Sci 6: 100–104. |
| [5] | Miller AJ, Smith SJ (1992) The mechanism of nitrate transport across the tonoplast of barley root cells. Planta 187: 554–557. |
| [6] | Jossier M, Kroniewicz L, Dalmas F, et al. (2010) The Arabidopsis vacuolar anion transporter, AtCLCc, is involved in the regulation of stomatal movements and contributes to salt tolerance. Plant J 4: 563–576. |
| [7] | Fischer-Schliebs E, Mariaux JB, Lüttge U (1997) Stimulation of H+transport activity of vacuolar H+ATPase by activation of H+PPase in Kalanchoë blossfeldiana. Biologia Plantarum 2: 169–177. |
| [8] | Taiz L (1992) The plant vacuole. J Exp Biol 172: 113–122. |
| [9] |
Schnitzer D, Seidel T, Sander T, et al. (2011) The cellular energization state affects peripheral stalk stability of plant vacuolar H+-ATPase and impairs vacuolar acidification. Plant Cell Physiol 52: 946–956. doi: 10.1093/pcp/pcr044
|
| [10] | Krebs M, Beyhl D, Gorlich E, et al. (2010) Arabidopsis V-ATPase activity at the tonoplast is required for efficient nutrient storage but not for sodium accumulation. Proc Nat Acad Sci USA 7: 3251–3256. |
| [11] | Smart LB, Vojdani F, Maeshima M, et al. (1998) Genes involved in osmoregulation during turgor-driven cell expansion of developing cotton fibers are differentially regulated. Plant Physiol 116: 1539–1549. |
| [12] |
Grabe M, Wang H, Oster G (2000) The mechanochemistry of V-ATPase proton pumps. Biophys J 78: 2798–2813. doi: 10.1016/S0006-3495(00)76823-8
|
| [13] |
Konishi H, Yamane H., Maeshima M, et al. (2004) Characterization of fructose-bisphosphate aldolase regulated by gibberellin in roots of rice seedlings. Plant Mol Biol 56: 839–848. doi: 10.1007/s11103-004-5920-2
|
| [14] |
Konishi H, Maeshima M, Komatsu S (2005) Characterization of vacuolar membrane proteins changed in rice root treated with gibberellin. J Proteome Res 4: 1775–1780. doi: 10.1021/pr050079c
|
| [15] | Klychnikov OI, Li KW, Lill H, et al. (2007) The V-ATPase from etiolated barley (Hordeum vulgare L) shoots is activated by blue light and interacts with 14-3-3 proteins. J Exp Bot 58: 1013–1023. |
| [16] |
Lu M, Ammar D, Ives H, et al. (2007) Physical interaction between aldolase and vacuolar H+-ATPase is essential for the assembly and activity of the proton pump. J Biol Chem 282: 24495–24503. doi: 10.1074/jbc.M702598200
|
| [17] | McCubbin AG, Ritchie SM, Swanson SJ, et al. (2004) The calcium-dependent protein kinase HvCDPK1 mediates the gibberellic acid response of the barley aleurone through regulation of vacuolar function. Plant J 39: 206–218. |
| [18] |
Hong-Hermesdorf A, Brüx A, Grüber A, et al. (2006) A WNK-kinase binds and phosphorylates V-ATP subunit C. FEBS Lett 580: 932–939. doi: 10.1016/j.febslet.2006.01.018
|
| [19] |
Cohen A, Perzov N, Nelson, et al. (1999) A novel family of yeast chaperons involved in the distribution of V-ATPase and other membrane proteins. J Biol Chem 274: 26885–26893. doi: 10.1074/jbc.274.38.26885
|
| [20] |
Holliday LS, Lu M, Lee BS, et al. (2000) The amino-terminal domain of the B subunit of vacuolar H+-ATPase contains a filamentous actin binding site. J Biol Chem 275: 32331–32337. doi: 10.1074/jbc.M004795200
|
| [21] |
Müller O, Neumann H, Bayer MJ, et al. (2003) Role of the Vtc proteins in V-ATPase stability and membrane trafficking. J Cell Sci 116: 1107–1115. doi: 10.1242/jcs.00328
|
| [22] |
Bayer MJ, Reese C, Buhler S, et al. (2003) Vacuole membrane fusion: V0 functions after trans-SNARE pairing and is coupled to the Ca2+-releasing channel. J Cell Biol 162:211–222. doi: 10.1083/jcb.200212004
|
| [23] |
Vitavska O, Merzendorfer H, Wieczorek H (2005) The V-ATpase subunit C binds to polymeric F-actin as well as to monomeric G-actin and induces crosslinking of actin filaments. J Biol Chem 280: 1070–1076. doi: 10.1074/jbc.M406797200
|
| [24] |
Cagnac O, Leterrier M, Yeager M, et al. (2007) Identification and characterization of Vxn1p, a novel type of vacuolar monovalent cation/H+-antiporter of Saccharomyces cerevisiae. J Biol Chem 282: 24284–24293. doi: 10.1074/jbc.M703116200
|
| [25] |
Smardon AM, Kane PM (2007) RAVE is essential for the efficient assembly of the C subunit with the vacuolar H(+)-ATPase. J Biol Chem 282:26185–26194. doi: 10.1074/jbc.M703627200
|
| [26] | Jones AM, Xuan Y, Xu M, et al. (2014) Border control- a membrane-linked interactome of Arabidopsis. Science 344: 711–716. |
| [27] |
Arabidopsis Interactome Mapping Consortium (2011) Evidence for network evolution in an Arabidopsis interactome map. Science 333: 601–607. doi: 10.1126/science.1203877
|
| [28] |
Trezzini GF, Horrichs A, Somssich IE (1993). Isolation of putative defenserelated genes from Arabidopsis thaliana and expression in fungal elicitor-treated cells. Plant Mol Biol 21: 385–389 doi: 10.1007/BF00019954
|
| [29] |
Alonso JM, Stepanova AN, Leisse TJ, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657. doi: 10.1126/science.1086391
|
| [30] |
Dietz KJ, Heber U, Mimura T (1998) Modulation of the vacuolar H+-ATPase by adenylates as basis for the transient CO2-dependent acidification of the leaf vacuole upon illumination. Biochim biophys acta 1373: 87–92. doi: 10.1016/S0005-2736(98)00094-7
|
| [31] |
Bencini DA, Shanley MS, Wild JR, et al. (1983) New assay for enzymatic phosphate release: application to aspartate transcarbamylase and other enzymes. Anal Biochem 132: 259–264. doi: 10.1016/0003-2697(83)90005-2
|
| [32] | Seidel T, Schnitzer D, Golldack D, et al. (2008) Organelle-specific isoenzymes of plant V-ATPae as revealed by in vivo-FRET analysis. BMC Cell Biol 9:928–928. |
| [33] |
Seidel T, Golldack D, Dietz KJ (2005) Mapping of C-termini of V-ATPase subunits by in vivo-FRET measurements. FEBS Lett 579: 4374–4382. doi: 10.1016/j.febslet.2005.06.077
|
| [34] | Müller SM, Galliardt H, Schneider J, et al. (2013) Quantification of Förster resonance energy transfer by monitoring sensitized emission in living plant cells. Front Plant Sci 4:413. |
| [35] | Obrdlik P, El.Bakkoury M, Hamacher T, et al. (2004) K+-channel interactions detected by a genetic system optimized for systematic studies of membrane protein interactions. Proc Nat Acad Sci USA 101: 12242–12247. |
| [36] |
Gietz RD, Schiestl RH, Willems AR, et al. (1995) Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast 11: 355–360. doi: 10.1002/yea.320110408
|
| [37] |
Ludewig U, Wilken S, Wu B et al. (2003) Homo- and hetero-oligomerization of ammonium transporter-1 NH4 uniporters. J Biol Chem 278: 45603–45610. doi: 10.1074/jbc.M307424200
|
| [38] |
Seidel T, Kluge C, Hanitzsch M, et al. (2004) Co-localization and FRET-analysis of subunits c and a of the vacuolar H+-ATPase in living plant cells. J Biotechnol 112: 165–175. doi: 10.1016/j.jbiotec.2004.04.027
|
| [39] | Appelhagen I, Huep G, Lu, GH, et al. (2010) Weird fingers: functional analysis of WIP domain proteins. FEBS Lett584: 3116–3122. |
| [40] |
Schwacke R, Schneider A, Van Der Graaff E, et al. (2003) ARAMEMNON, a Novel Database for Arabidopsis Integral Membrane Proteins. Plant Physiol 131: 16–26. doi: 10.1104/pp.011577
|
| [41] |
Obayashi T, Nishida K, Kasahara K, et al. (2011) ATTED-II updates: condition-specific gene coexpression to extend coexpression analyses and applications to a broad range of flowering plants. Plant Cell Physiol 52: 213–219. doi: 10.1093/pcp/pcq203
|
| [42] |
Joshi HJ, Hirsch-Hoffmann M, Baerenfaller K, et al. (2011) MASCP Gator: An aggregation portal for the visualization of Arabidopsis proteomics data. Plant Physiol 155: 259–270. doi: 10.1104/pp.110.168195
|
| [43] |
Winter D, Vinegar B, Ammar R, et al. (2007) An 'Electronic Fluorescent Pictograph' Browser for Exploring and Analyzing Large-Scale Biological Data Sets. PLoS One 2: e718. doi: 10.1371/journal.pone.0000718
|
| [44] |
Mäser P, Thomine S, Schroeder JI, et al. (2001) Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol 126: 1646–1667. doi: 10.1104/pp.126.4.1646
|
| [45] |
Cheng NH (2005). Functional Association of Arabidopsis CAX1 and CAX3 Is Required for Normal Growth and Ion Homeostasis. Plant Physiol 138: 2048–2060. doi: 10.1104/pp.105.061218
|
| [46] | Silva P, Gerós H (2014) Regulation by salt of vacuolar H+ -ATPase and H+ -pyrophosphatase activities and Na+ /H+ exchange. Plant Signal Behav 4: 718–726. |
| [47] |
Gadella T, Krogt G, Bisseling T (1999) GFP-based FRET microscopy in living plant cells. Trends Plant Sci 4: 287–291. doi: 10.1016/S1360-1385(99)01426-0
|
| [48] | Li J, Yang H, Peer WA, et al. (2005) Arabidopsis H+-PPase AVP1 regulates auxin-mediated organ development. Science 310: 121–125. |
| [49] | Viotti C (2014) ER and vacuoles: never been closer. Frontiers Plant Sci 5:20. |
| [50] |
Yoshida K, Miki N, Momonoi K, et al. (2009) Synchrony between flower opening and petal-color change from red to blue in morning glory, Ipomoea tricolor cv. Heavenly Blue. Proc Japan Acad Series B, Physic biol sci 85: 187–197. doi: 10.2183/pjab.85.187
|
| [51] |
Ali V, Shigeta Y, Tokumoto U, et al. (2004) An intestinalparasitic protist, Entamoeba histolytica, possesses a non-redundant nitrogen fixation-like system for iron-sulfur cluster assembly under anaerobic conditions. J Biol Chem 279: 16863–16874. doi: 10.1074/jbc.M313314200
|
| [52] | Gaxiola RA, Rao R, Sherman A, et al. (1999) The Arabidopsis thaliana proton transporters, AtNhx1 and Avp1, can function in cation detoxification in yeast. Proc Nat Acad Sci USA 96: 1480–1485. |
| [53] | Zhang Y, Wang L, Liu Y, et al. (2006. Nitric oxide enhances salt tolerance in maize seedlings through increasing activities of proton-pump and Na+/H+ antiport in the tonoplast. Planta 224: 545–555. |
| [54] | Bassil E, Tajima H, Liang YC, et al. (2011). The Arabidopsis Na+/H+antiporters NHX1 and NHX2 control vacuolar pH and K+ homeostasis to regulate growth, flower development, and reproduction. Plant Cell 23: 3482–3497. |
| [55] | Hirschi KD, Zhen RG, Cunningham KW, et al. (1996) CAX1, an H+/Ca2+ antiporter from Arabidopsis. Proc Nat Acad Sci USA 16: 8782–8786. |
| [56] |
Shigaki T, Pittman JK, Hirschi KD (2003) Manganese specificity determinants in the Arabidopsis metal/H+ antiporter CAX2. J Biol Chem 278: 6610–6617. doi: 10.1074/jbc.M209952200
|
| [57] |
Cheng NH (2003) The Arabidopsis cax1 Mutant Exhibits Impaired Ion Homeostasis, Development, and Hormonal Responses and Reveals Interplay among Vacuolar Transporters. Plant Cell 15: 347–364. doi: 10.1105/tpc.007385
|
| [58] | Barkla BJ, Hirschi KD, Pittman JK (2014) Exchangers man the pumps. Plant Signal Behav 3: 354–356. |
| [59] | Guern J, Mathieu Y, Kurkdjian A, et al. (1989) Regulation of Vacuolar pH of Plant Cells II. A 31P NMR Study of the Modifications of Vacuolar pH in Isolated Vacuoles Induced by Proton Pumping and Cation/H+ Exchanges. Plant Physiol 89: 27–36. |
| [60] |
Manohar M, Shigaki T, Hirschi KD (2011) Plant cation/H+ exchangers (CAXs): Biological functions and genetic manipulations. Plant Biol 13: 561–569. doi: 10.1111/j.1438-8677.2011.00466.x
|
| [61] |
Conn SJ, Gilliham M, Athman A, et al. (2011) Cell-specific vacuolar calcium storage mediated by CAX1 regulates apoplastic calcium concentration, gas exchange, and plant productivity in Arabidopsis. Plant Cell 23: 240–257. doi: 10.1105/tpc.109.072769
|
| [62] |
McIntyre GI (1997) The role of nitrate in the osmotic and nutritional control of plant development. Austral J Plant Physiol 24: 103–118. doi: 10.1071/PP96064
|
| [63] | Randall SK, Sze H (1986) Properties of the partially purified tonoplast H+-pumping ATPase from oat roots. J Biol Chem 261: 1364–1371. |
| [64] |
De Angeli A, Monachello D, Ephritikhine G, et al. (2006) The nitrate/proton antiporter AtCLCa mediates nitrate accumulation in plant vacuoles. Nature 442: 939–942. doi: 10.1038/nature05013
|
| [65] |
Fecht-Bartenbach J, Bogner M, Dynowski M, et al. (2010) CLCb-mediated NO-3/H+ exchange across the tonoplast of Arabidopsis vacuoles. Plant Cell Physiol 51: 960–968. doi: 10.1093/pcp/pcq062
|
| [66] |
Wudick MM, Luu DT, Maurel C (2009) A look inside: localization patterns and functions of intracellular plant aquaporins. New phytol 184: 289–302. doi: 10.1111/j.1469-8137.2009.02985.x
|
| [67] |
Geelen D, Lurin C, Bouchez D, et al. (2000). Disruption of putative anion channel gene AtCLC-a in Arabidopsis suggests a role in the regulation of nitrate content. Plant J Cell Mol Biol 21: 259–267. doi: 10.1046/j.1365-313x.2000.00680.x
|
| [68] |
Harada H, Kuromori T, Hirayama T, et al. (2004) Quantitative trait loci analysis of nitrate storage in Arabidopsis leading to an investigation of the contribution of the anion channel gene, AtCLC-c, to variation in nitrate levels. J Exp Bot 55: 2005–2014. doi: 10.1093/jxb/erh224
|
| [69] |
Miller AJ, Fan X, Orsel M, et al. (2007) Nitrate transport and signalling. J Exp Bot 58: 2297–2306. doi: 10.1093/jxb/erm066
|
| [70] |
Fecht-Bartenbach J, Bogner M, Krebs M, et al. (2007) Function of the anion transporter AtCLC-d in the trans-Golgi network. Plant J 50: 466–474. doi: 10.1111/j.1365-313X.2007.03061.x
|
| [71] |
Wilkinson JQ, Crawford NM (1991) Identification of the Arabidopsis CHL3 gene as the nitrate reductase structural gene NIA2. Plant Cell 3: 461–471. doi: 10.1105/tpc.3.5.461
|
| [72] |
Trewava A (1983) Plant growth substances—Metabolic flywheels for plant development. Cell Biol Intern Rep 7: 569–575. doi: 10.1016/0309-1651(83)90109-1
|
| [73] |
Kaiser WM, Weiner H, Huber SC (1999) Nitrate reductase in higher plants: A case study for transduction of environmental stimuli into control of catalytic activity. Physiologia plantarum 105: 384–389. doi: 10.1034/j.1399-3054.1999.105225.x
|
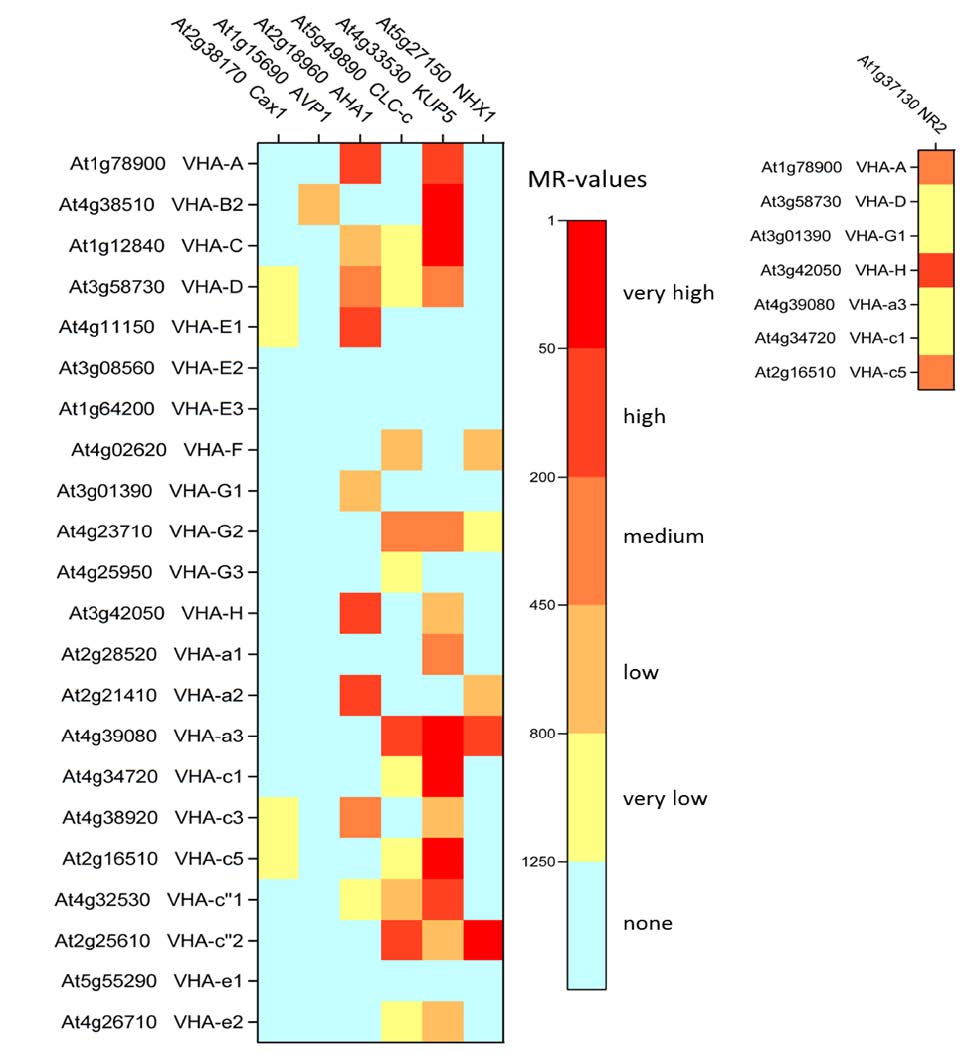
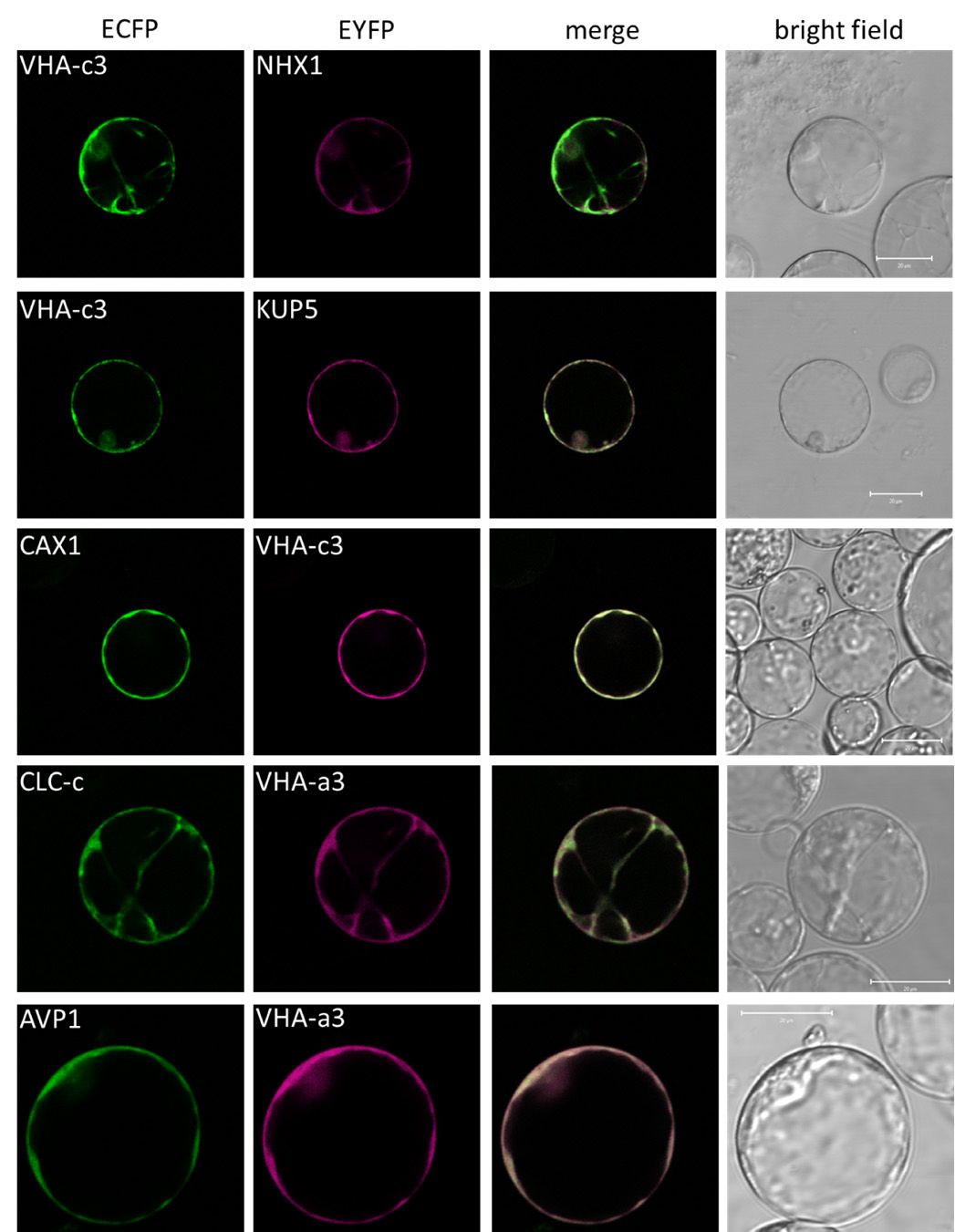
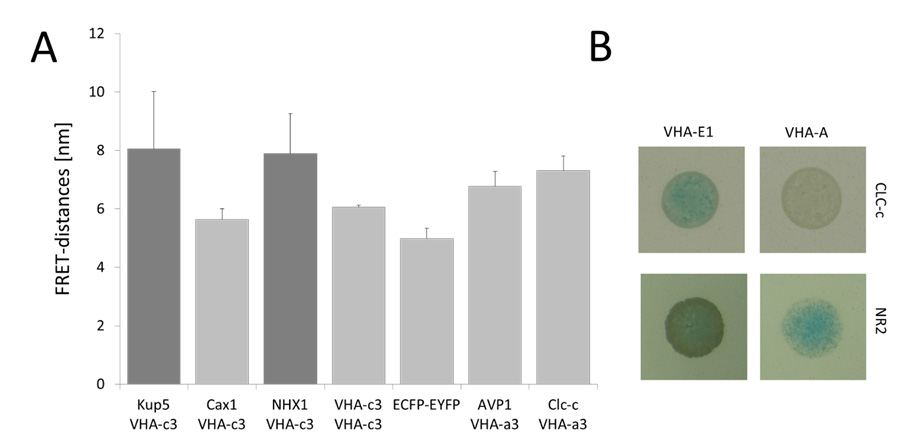

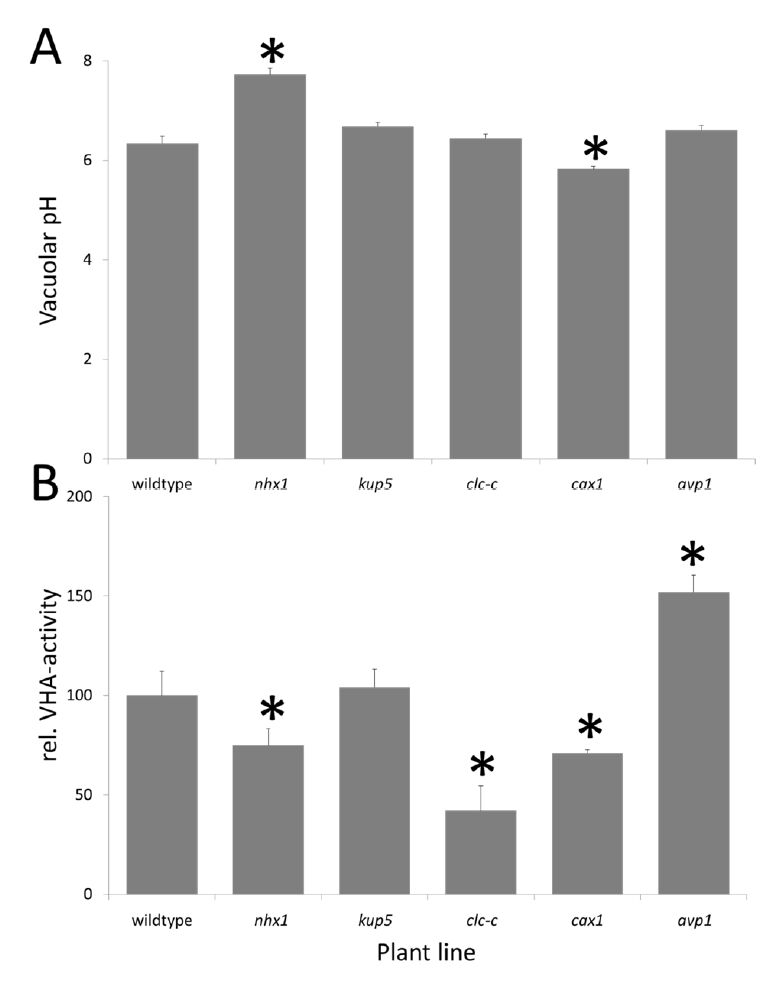
Figures(5) / Tables(3)

Michèle Siek, Berenice Marg, Chris M. Ehring, Derya Kirasi, Michael Liebthal, Thorsten Seidel. Interplay of vacuolar transporters for coupling primary and secondary active transport[J]. AIMS Biophysics, 2016, 3(4): 479-500. doi: 10.3934/biophy.2016.4.479







 DownLoad:
DownLoad: