Citation: Alexander G. Volkov, Yuri B. Shtessel. Propagation of electrotonic potentials in plants: Experimental study and mathematical modeling[J]. AIMS Biophysics, 2016, 3(3): 358-379. doi: 10.3934/biophy.2016.3.358

| [1] | Herde O, Peña-Cortés H, Fisahn J (1995) Proteinase inhibitor II gene expression induced by electrical stimulation and control of photosynthetic activity in tomato plants. Plant Cell Physiol 36: 737–742. |
| [2] | Wildon DC, Thain JF, Minchin PEH, et al. (1992) Electric signaling and systemic proteinase inhibitor induction in the wounded plant. Nature 360: 62–65. |
| [3] |
Stankovic B, Davies E (1997) Intercellular communication in plants: electrical stimulation of proteinase inhibitor gene expression in tomato. Planta 202: 402–406. doi: 10.1007/s004250050143

|
| [4] | Inaba A, Manabe T, Tsuji H, et al. (1995) Electrical impedance analysis of tissue properties associated with ethylene induction by electric currents in cucumber (Cucumis sativus L.) fruit. J Plant Physiol 107: 199–205. |
| [5] | Favre P, Agosti RD (2007) Voltage-dependent action potentials in Arabidopsis thaliana. Physiol Plantarum 131: 263–272. |
| [6] | Volkov AG (Ed.) (2012) Plant Electrophysiology. Methods and Cell Electrophysiology. Berlin: Springer. |
| [7] | Volkov AG (Ed.) (2012) Plant Electrophysiology. Signaling and Responses. Berlin: Springer. |
| [8] |
Balmer RT, Franks JG (1975) Contractile characteristics of Mimosa pudica L. J Plant Physiol 56: 464–467. doi: 10.1104/pp.56.4.464
|
| [9] |
Jonas H (1970) Oscillations and movements of Mimosa leaves due to electric shock. J Interdiscipl Cycle 1: 335–348. doi: 10.1080/09291017009359229
|
| [10] | Volkov AG (2016) Biosensors, memristors and actuators in electrical networks of plants. Intern J Parallel Emerg Distrib Systems 1–12. |
| [11] |
Volkov AG, Adesina T, Jovanov E (2008) Charge induced closing of Dionaea muscipula Ellis trap. Bioelectrochem 74: 16–21. doi: 10.1016/j.bioelechem.2008.02.004
|
| [12] | Volkov AG, Adesina T, Markin VS, et al. (2008) Kinetics and mechanism of Dionaea muscipula trap closing. Plant Physiol 146: 694–702. |
| [13] |
Volkov AG, Coopwood KJ, Markin VS (2008) Inhibition of the Dionaea muscipula Ellis trap closure by ion and water channels blockers and uncouplers. Plant Sci 175: 642–649. doi: 10.1016/j.plantsci.2008.06.016
|
| [14] |
Volkov AG, Carrell H, Baldwin A, et al. (2009) Electrical memory in Venus flytrap. Bioelectrochem 75: 142–147. doi: 10.1016/j.bioelechem.2009.03.005
|
| [15] |
Volkov AG, Carrell H, Markin VS (2009) Biologically closed electrical circuits in Venus flytrap. Plant Physiol 149: 1661–1667. doi: 10.1104/pp.108.134536
|
| [16] |
Volkov AG, Foster JC, Ashby TA, et al. (2010) Mimosa pudica: electrical and mechanical stimulation of plant movements. Plant Cell Environ 33: 163–173. doi: 10.1111/j.1365-3040.2009.02066.x
|
| [17] |
Black J, Forsyth F, Fensom D, et al. (1971) Electrical stimulation and its effects on growth and ion accumulation in tomato plants. Can J Bot 49: 1809–1815. doi: 10.1139/b71-255
|
| [18] | Gensler W (1974) Bioelectric potential and their relation to growth in higher plants. Ann NY Acad Sci 238: 281–299. |
| [19] | Murr L.E (1963) Plant growth response in a simulated electric field environment. Nature 200: 490–491. |
| [20] | Takamura T (2006) Electrochemical potential around the plant root in relation to metabolism and growth acceleration, in: Volkov AG (ed), Plant Electrophysiology – Theory & Methods. Berlin: Springer, 341–374. |
| [21] |
Markin VS, Volkov AG, Chua L (2014) An analytical model of memristors in plants. Plant Signal Behav 9: e972887-1-9. doi: 10.4161/15592316.2014.972887
|
| [22] |
Volkov AG, Forde-Tuckett V, Reedus J, et al. (2014) Memristor in the Venus flytrap. Plant Signal Behav 9: e29204-1-12. doi: 10.4161/psb.29204
|
| [23] |
Volkov AG, Reedus J, Mitchell CM, et al. (2014) Memristor in the electrical network of Aloe vera L. Plant Signal Behav 9: e29056-1-7. doi: 10.4161/psb.29056
|
| [24] |
Volkov AG, Reedus J, Mitchell CM, et al. (2014) Memory elements in the electrical network of Mimosa pudica L. Plant Signal Behav 9: e982029-1-9. doi: 10.4161/15592324.2014.982029
|
| [25] |
Volkov AG, Tuckett C, Reedus J, et al. (2014) Memristors in plants. Plant Signal Behav 9: e28152-1-8. doi: 10.4161/psb.28152
|
| [26] |
Volkov AG, Nyasani EK, Blackmon AL, et al. (2015) Memristors: Memory elements in potato tubers. Plant Signal Behav 10: e1071750-1-7. doi: 10.1080/15592324.2015.1071750
|
| [27] | Volkov AG, Nyasani EK, Tuckett C, et al. (2016). Electrophysiology of Pumpkin Seeds: Memristors in Vivo. Plant Signal Behav 11: e1151600; DOI: 10.1080/15592324.2016.1151600. |
| [28] |
Frachisse-Stoilskovic JM, Julien JL (1993) The coupling between extra-and intracellular electric potentials in Bidens pilosa L. Plant Cell Environm 16: 633–641. doi: 10.1111/j.1365-3040.1993.tb00481.x
|
| [29] | Overall RL, Gunning BES (1982) Intercellular communication in Azolla roots: II. Electrical coupling. Protoplasma 111: 151–160. |
| [30] | Sibaoka T, Tabata T (1981) Electrotonic coupling between adjacent internodal cells of Chara: transmission of action potentials beyond the node. Plant Cell Physiol 22: 397–411. |
| [31] |
Spanswick RM (1972) Electrical coupling between cells of higher plants: A direct demonstration of intercellular communication. Planta 102: 215–227. doi: 10.1007/BF00386892
|
| [32] | Volkov AG, O’Neal L, Volkova-Gugeshashvili MI, et al. (2013) Electrostimulation of Aloe Vera L., Mimosa Pudica L. and Arabidopsis Thaliana: Propagation and collision of electronic potentials. J Electrochem Soc 160: G3102–G3111. |
| [33] | Volkov AG, Vilfranc CL, Murphy VA, et al. Electrotonic and action potentials in the Venus flytrap. J Plant Physiol 170: 838–846. |
| [34] | Jack JJ, Noble D, Tsien RW (1975) Electric current flow in excitable cells. Clarendon, Oxford. |
| [35] | Adamatzky A (2014) Towards plant wires. Biosystems 122: 1–6. |
| [36] |
Stavrinidou E, Gabrielsson R, Gomez E, et al. (2015) Electronic plants. Sci Adv 1: e1501136-1-8. doi: 10.1126/sciadv.1501136
|
| [37] |
Hodgkin AL, Rushton WAH (1946) The electrical constants of a crustacean nerve fibre. Proc Royal Soc B 133: 444–479. doi: 10.1098/rspb.1946.0024
|
| [38] | Rall W (1969) Time constants and electrotonic length of membrane cylinders and neurons. Biophys J 58: 631–639. |
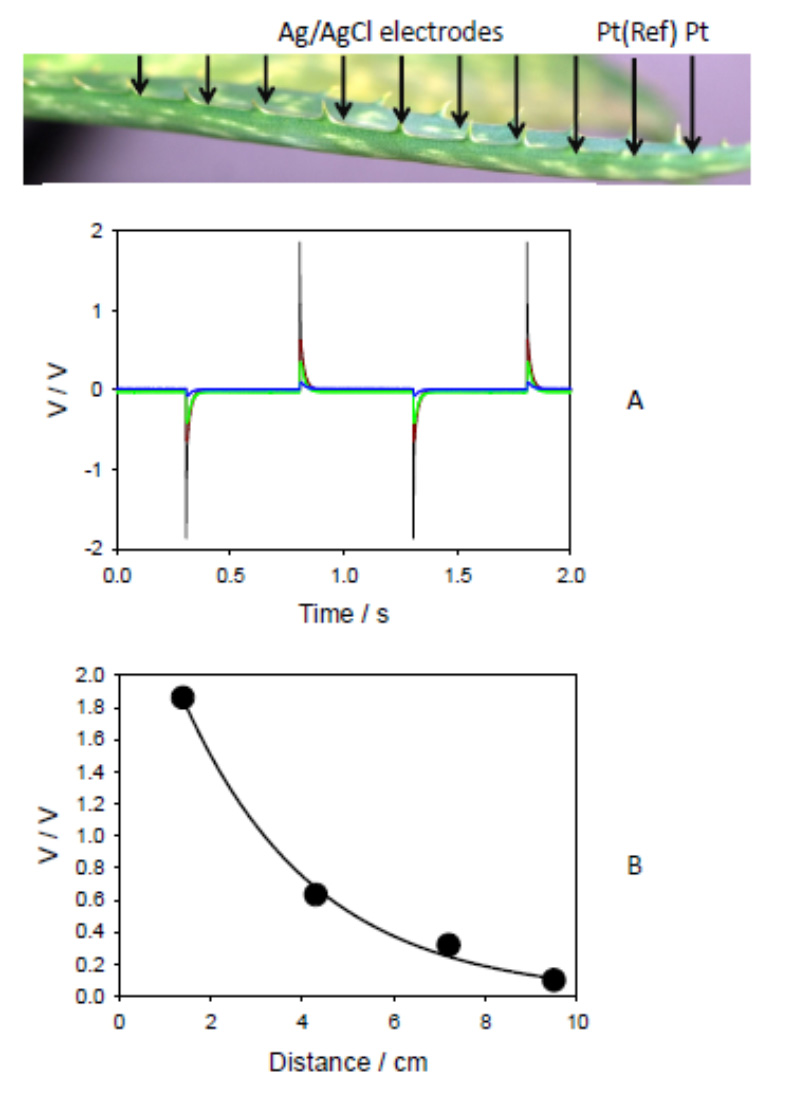
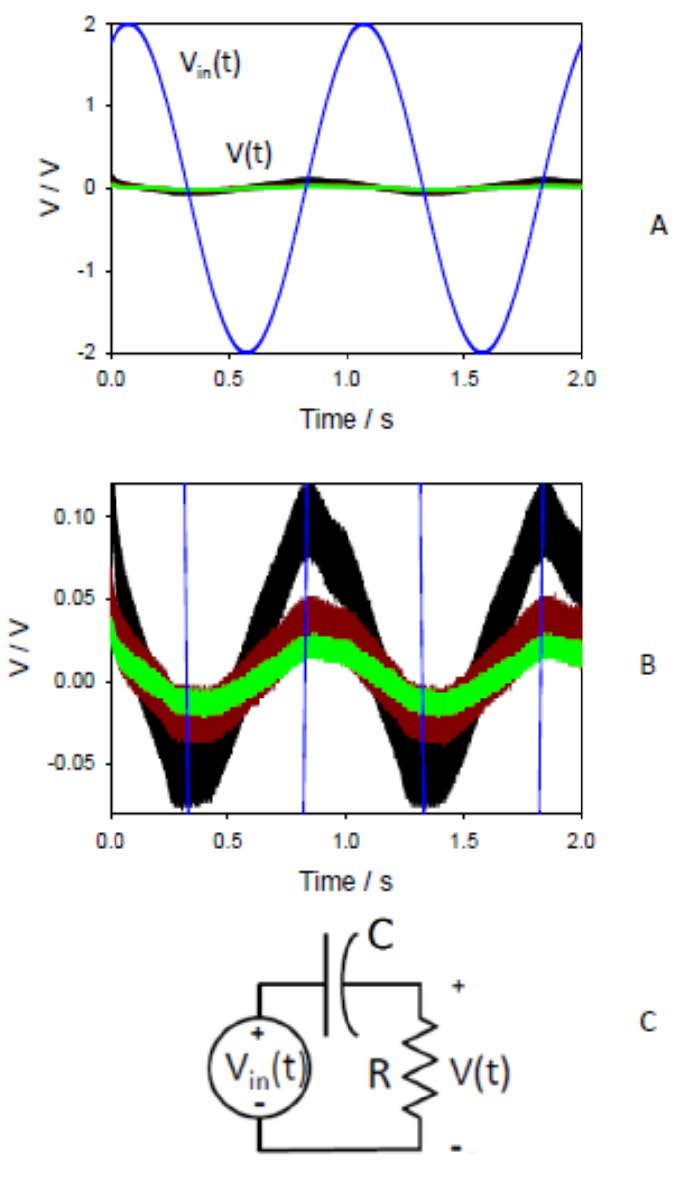
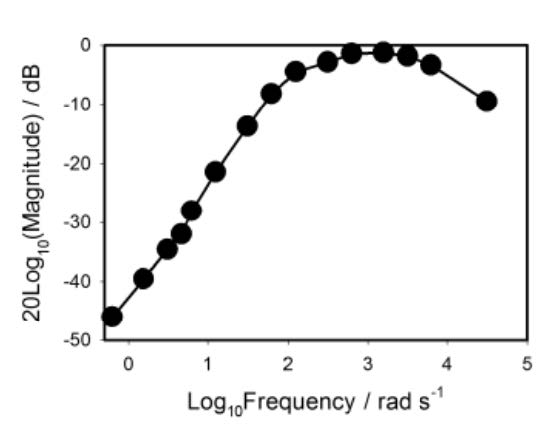
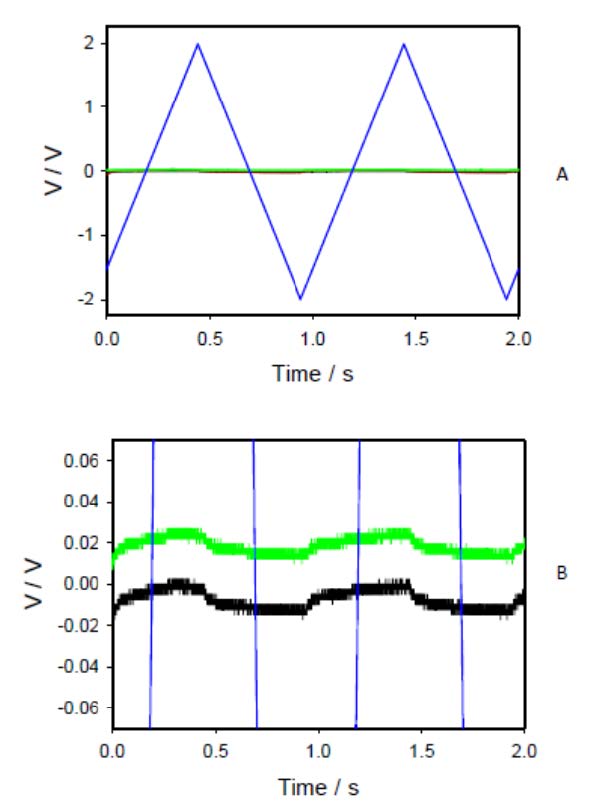
Figures(13)

Alexander G. Volkov, Yuri B. Shtessel. Propagation of electrotonic potentials in plants: Experimental study and mathematical modeling[J]. AIMS Biophysics, 2016, 3(3): 358-379. doi: 10.3934/biophy.2016.3.358







 DownLoad:
DownLoad: