Citation: Hanieh Hadady, Doug Redelman, Sage R. Hiibel, Emil J. Geiger. Continuous-flow sorting of microalgae cells based on lipid content by high frequency dielectrophoresis[J]. AIMS Biophysics, 2016, 3(3): 398-414. doi: 10.3934/biophy.2016.3.398

| [1] |
Hu Q, Sommerfeld M, Jarvis E, et al. (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J 54: 621–639. doi: 10.1111/j.1365-313X.2008.03492.x

|
| [2] |
Chisti Y (2008) Biodiesel from microalgae beats bioethanol. Trends Biotechnol 26: 126–131. doi: 10.1016/j.tibtech.2007.12.002
|
| [3] |
Dismukes GC, Carrieri D, Bennette N, et al. (2008) Aquatic phototrophs: efficient alternatives to land-based crops for biofuels. Curr Opin Biotech 19: 235–240. doi: 10.1016/j.copbio.2008.05.007
|
| [4] | Sharif H, Salleh A, Boyce AN, et al. Biodiesel Fuel Production from Algae as Renewable Energy. Am J Bioch Biotechnol 2008 4: 250–254. |
| [5] |
Hannon M, Gimpel J, Tran M, et al. (2010) Biofuels from algae: challenges and potential. Curr Opin Biotech 21: 277–286. doi: 10.1016/j.copbio.2010.03.005
|
| [6] |
Bono MS, Ahner BA, Kirby BJ (2013) Detection of algal lipid accumulation due to nitrogen limitation via dielectric spectroscopy of Chlamydomonas reinhardtii suspensions in a coaxial transmission line sample cell. Bioresource Technol 143: 623–631. doi: 10.1016/j.biortech.2013.06.040
|
| [7] |
Deng YL, Kuo MY, Juang YJ (2014) Development of flow through dielectrophoresis microfluidic chips for biofuel production: Sorting and detection of microalgae with different lipid contents. Biomicrofluidics 8: 064120–064120. doi: 10.1063/1.4903942
|
| [8] |
Hadady H, Montiel C, Wetta D, et al. (2015) Liposomes as a model for the study of high frequency dielectrophoresis. Electrophoresis 36: 1423–1428. doi: 10.1002/elps.201400480
|
| [9] |
Hadady H, Wong JJ, Hiibel SR, et al. (2014) High frequency dielectrophoretic response of microalgae over time. Electrophoresis 35: 3533–3540. doi: 10.1002/elps.201400306
|
| [10] |
Michael KA, Hiibel SR, Geiger EJ (2014) Dependence of the dielectrophoretic upper crossover frequency on the lipid content of microalgal cells. Algal Research 6: 17–21. doi: 10.1016/j.algal.2014.08.004
|
| [11] |
Gagnon ZR (2011) Cellular dielectrophoresis: applications to the characterization, manipulation, separation and patterning of cells. Electrophoresis 32: 2466–2487. doi: 10.1002/elps.201100060
|
| [12] | Hoettges KF (2009) Dielectrophoresis as a Cell Characterisation Tool. Microengineering in Biotechnology, Meth Mol Biol. New York. NY: Humana Press, 183–198. |
| [13] | Hughes MP (2002) Strategies for dielectrophoretic separation in laboratory-on-a-chip systems. Electrophoresis 23: 2569–2582. |
| [14] |
Gagnon Z, Gordon J, Sengupta S, et al. (2008) Bovine red blood cell starvation age discrimination through a glutaraldehyde-amplified dielectrophoretic approach with buffer selection and membrane cross-linking. Electrophoresis 29: 2272–2279. doi: 10.1002/elps.200700604
|
| [15] |
Song H, Rosano JM, Wang Y, et al. (2015) Continuous-flow sorting of stem cells and differentiation products based on dielectrophoresis. Lab Chip 15: 1320–1328. doi: 10.1039/C4LC01253D
|
| [16] | Valero A, Braschler T, Demierre N, et al. (2010) A miniaturized continuous dielectrophoretic cell sorter and its applications. Biomicrofluidics 4: 1–22. |
| [17] |
Gossett DR, Weaver WM, Mach AJ, et al. (2010) Label-free cell separation and sorting in microfluidic systems. Anal and Bioanal Chem 397: 3249–3267. doi: 10.1007/s00216-010-3721-9
|
| [18] |
Mernier G, Piacentini N, Braschler T, et al. (2010) Continuous-flow electrical lysis device with integrated control by dielectrophoretic cell sorting. Lab Chip 10: 2077–2082. doi: 10.1039/c000977f
|
| [19] |
Vahey MD, Voldman J (2008) An Equilibrium Method for Continuous-Flow Cell Sorting Using Dielectrophoresis. Anal Chem 80: 3135–3143. doi: 10.1021/ac7020568
|
| [20] |
Hu X, Bessette PH, Qian J, et al. (2005) Marker-specific sorting of rare cells using dielectrophoresis. Pnas 102: 15757–15761. doi: 10.1073/pnas.0507719102
|
| [21] |
Wang X-B, Yang J, Huang Y, et al. (2000) Cell Separation by Dielectrophoretic Field-flow-fractionation. Anal Chem 72: 832–839. doi: 10.1021/ac990922o
|
| [22] |
Moon HS, Kwon K, Kim SI, et al. (2011) Continuous separation of breast cancer cells from blood samples using multi-orifice flow fractionation (MOFF) and dielectrophoresis (DEP). Lab Chip 11: 1118–1125. doi: 10.1039/c0lc00345j
|
| [23] |
Doh I, Cho YH (2005) A continuous cell separation chip using hydrodynamic dielectrophoresis (DEP) process. Sensor Actuat A-Phys 121: 59–65. doi: 10.1016/j.sna.2005.01.030
|
| [24] | Yang J, Huang Y, Wang XB, et al. (1999) Cell Separation on Microfabricated Electrodes Using Dielectrophoretic/Gravitational Field-Flow Fractionation. J Am Chem Soc 71: 911–918. |
| [25] | Lewpiriyawong N, Yang C (2013) Dielectrophoresis Field-Flow Fractionation for Continuous-Flow Separation of Particles and Cells in Microfluidic Devices. Advances in Transport Phenomena 2011. Switzerland: Springer International Publishing, 29–62. |
| [26] |
Pommer MS, Zhang Y, Keerthi N, et al. (2008) Dielectrophoretic separation of platelets from diluted whole blood in microfluidic channels. Electrophoresis 29: 1213–1218. doi: 10.1002/elps.200700607
|
| [27] |
Cheng IF, Chang HC, Hou D (2007) An integrated dielectrophoretic chip for continuous bioparticle filtering, focusing, sorting, trapping, and detecting. Biomicrofluidics 1: 21503–21515. doi: 10.1063/1.2723669
|
| [28] |
Piacentini N, Mernier G, Tornay R, et al. (2011) Separation of platelets from other blood cells in continuous-flow by dielectrophoresis field-flow-fractionation. Biomicrofluidics 5: 034122–034128. doi: 10.1063/1.3640045
|
| [29] |
Cima I, Yee CW, Iliescu FS, et al. (2013) Label-free isolation of circulating tumor cells in microfluidic devices: Current research and perspectives. Biomicrofluidics 7: 011810–011813. doi: 10.1063/1.4780062
|
| [30] |
Deng YL, Chang JS, Juang YJ (2013) Separation of microalgae with different lipid contents by dielectrophoresis. Bioresource Technol 135: 137–141. doi: 10.1016/j.biortech.2012.11.046
|
| [31] | Schor AR, Buie CR (2012) Non-Invasive Sorting of Lipid Producing Microalgae With Dielectrophoresis Using Microelectrodes. ASME 2012 International Mechanical Engineering Congress and Exposition. Houston, Texas: American Society of Mechanical Engineers, 701–707. |
| [32] | Schor AR, Buie CR (2015) Dielectrophoretic sorting of lipid-containing microorganisms using high frequency electric fields produced by conducting post arrays. Solid-State Sensors, Actuators and Microsystems (TRANSDUCERS), 2015 Transducers - 2015 18th International Conference on. Anchorage, AK: IEEE, 1617–1620. |
| [33] |
Broche LM, Labeed FH, Hughes MP (2005) Extraction of dielectric properties of multiple populations from dielectrophoretic collection spectrum data. Phys Med Biol 50: 2267–2274. doi: 10.1088/0031-9155/50/10/006
|
| [34] |
Han KH, Frazier AB (2008) Lateral-driven continuous dielectrophoretic microseparators for blood cells suspended in a highly conductive medium. Lab Chip 8: 1079–1086. doi: 10.1039/b802321b
|
| [35] |
Shima HC, Kwaka YK, Hanb CS, et al. (2009) Effect of a square wave on an assembly of multi-walled carbon nanotubes using AC dielectrophoresis. Physica E: Low-dimensional Systems and Nanostructures 41: 1137–1142. doi: 10.1016/j.physe.2008.12.007
|
| [36] | Rodolfi L, Chini Zittelli G, Padovani G, et al. (2014) Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnol Bioeng 102: 100–112. |
| [37] |
Li X, Moellering ER, Liu B, et al. (2012) A Galactoglycerolipid Lipase Is Required for Triacylglycerol Accumulation and Survival Following Nitrogen Deprivation in Chlamydomonas reinhardtii. The Plant Cell 24: 4670–4686. doi: 10.1105/tpc.112.105106
|
| [38] | Sheehan J, Dunahay T, Benemann J, et al. (1998) Look back at the U. S. Department of Energy's Aquatic Species Program: Biodiesel from Algae. U.S. Department of Energy’s Office of Fuels Development. |
| [39] | Hadady H, Wong JJ, Hiibel SR, et al. (2014) Effect of Media Conductivity on High Frequency Dielectrophoretic Response. ASME 2014 International Mechanical Engineering Congress and Exposition. Montreal, Quebec, Canada: American Society of Mechanical Engineers. V010T013A006. |
| [40] |
Hoettges KF, Hubner Y, Broche LM, et al. (2008) Dielectrophoresis-activated multiwell plate for label-free high-throughput drug assessment. Anal Chem 80: 2063–2068. doi: 10.1021/ac702083g
|
| [41] |
Cooper MS, Hardin WR, Petersen TW, et al. (2010) Visualizing "green oil" in live algal cells. J Biosci Bioeng 109: 198–201. doi: 10.1016/j.jbiosc.2009.08.004
|
| [42] | Akimoto S, Mimuro M (2007) Application of Time‐Resolved Polarization Fluorescence Spectroscopy in the Femtosecond Range to Photosynthetic Systems. J Photoch Photobio 83: 163–170. |
| [43] | Xia Y, Whitesides GM (2003) SOFT LITHOGRAPHY. Annu Rev Mater Sci 28: 159–184. |
| [44] |
Geiger EJ, Pisano AP, Svec F (2010) A Polymer-Based Microfluidic Platform Featuring On-Chip Actuated Hydrogel Valves for Disposable Applications. J Microelectromech S 19: 944–950. doi: 10.1109/JMEMS.2010.2048702
|
| [45] |
Aran K, Sasso LA, Kamdar N, et al. (2010) Irreversible, direct bonding of nanoporous polymer membranes to PDMS or glass microdevices. Lab Chip 10: 548–552. doi: 10.1039/b924816a
|
| [46] |
Gesche R, Kovacs R, Scherer J (2005) Mobile plasma activation of polymers using the plasma gun. Surf Coat Tech 200: 544–547. doi: 10.1016/j.surfcoat.2005.01.109
|
| [47] | Hadady H, Michael KA, Geiger EJ (2014) Impedance Effects During High–Frequency Dielectrophoresis. ASME 2014 International Mechanical Engineering Congress and Exposition. Montreal, Quebec, Canada: American Society of Mechanical Engineers, V010T013A046. |
| [48] |
Mooij PR, Stouten GR, Tamis J, et al. (2013) Survival of the fattest. Energ Environ Sci 6: 3404–3406. doi: 10.1039/c3ee42912a
|
| [49] |
Ling SH, Lam YC, Chian KS (2012) Continuous Cell Separation Using Dielectrophoresis through Asymmetric and Periodic Microelectrode Array. Anal Chem 84: 6463–6470. doi: 10.1021/ac300079q
|
| [50] | Hadady H, Hiibel SR, Redelman D, et al. (2015) Use of a Separability parameter for the Design of a High Frequency Dielectrophoresis Cell Sorter Device. InterPACK/ICNMM2015. California. |
| [51] | Bono MS, Garcia RD, Sri-Jayantha DV, et al. (2015) Measurement of Lipid Accumulation in Chlorella vulgaris via Flow Cytometry and Liquid-State ?1H NMR Spectroscopy for Development of an NMR-Traceable Flow Cytometry Protocol. Plos One 10: 1–18. |
| [52] | Hyka P, Lickova S, P?ibyl P, et al. (2013) Flow cytometry for the development of biotechnological processes with microalgae. Biotechnol Adv 31: 2–16. |
| [53] |
Montero MF, Aristizábal M, Reina GG (2011) Isolation of high-lipid content strains of the marine microalga Tetras. J Appl Phycol 23: 1053–1057. doi: 10.1007/s10811-010-9623-6
|
| [54] |
Velmurugan N, Sung M, Yim SS, et al. (2013) Evaluation of intracellular lipid bodies in Chlamydomonas reinhardtii strains by flow cytometry. Bioresource Technol 138: 30–37. doi: 10.1016/j.biortech.2013.03.078
|
| [55] |
Wang ZT, Ullrich N, Joo S, et al. (2009) Algal Lipid Bodies: Stress Induction, Purification, and Biochemical Characterization in Wild-Type and Starchless Chlamydomonas reinhardtii. Eukaryot Cell 8: 1856–1868. doi: 10.1128/EC.00272-09
|
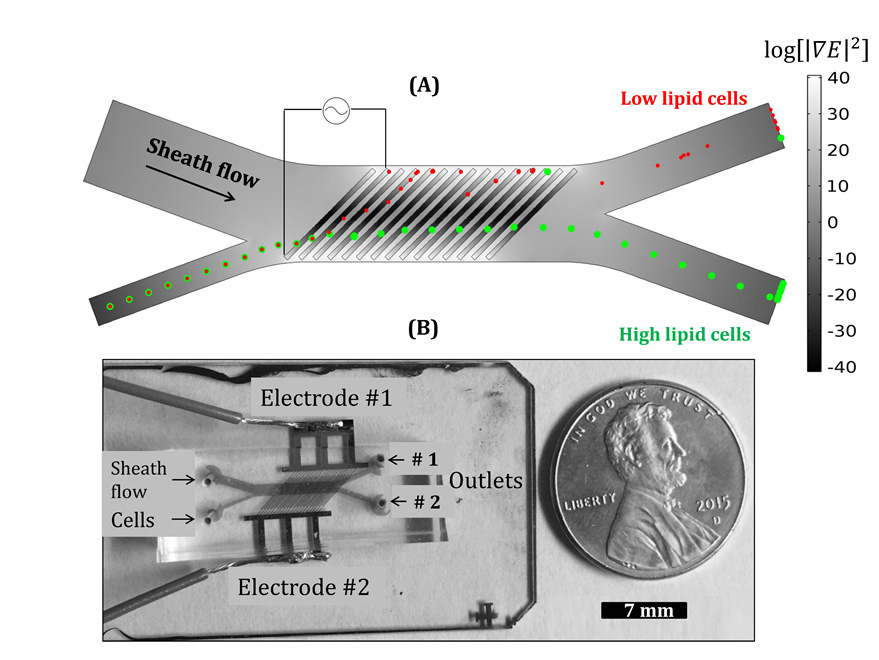
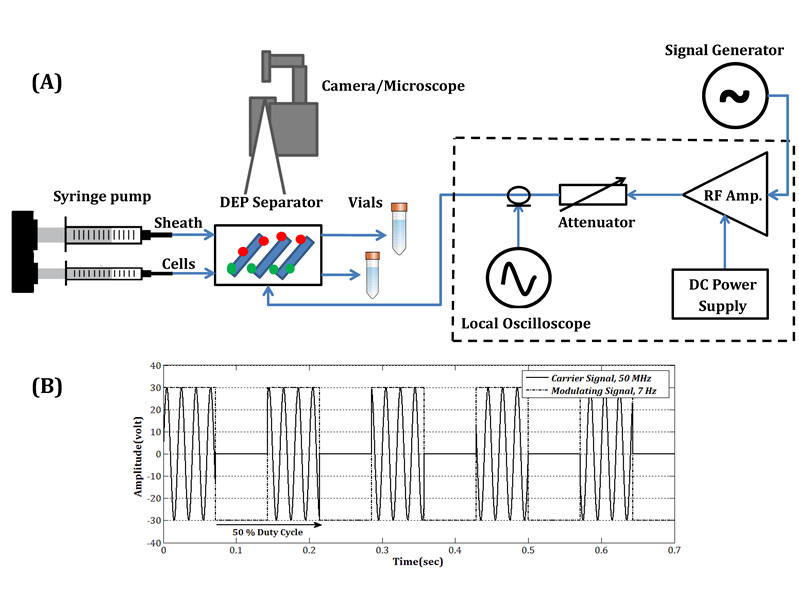


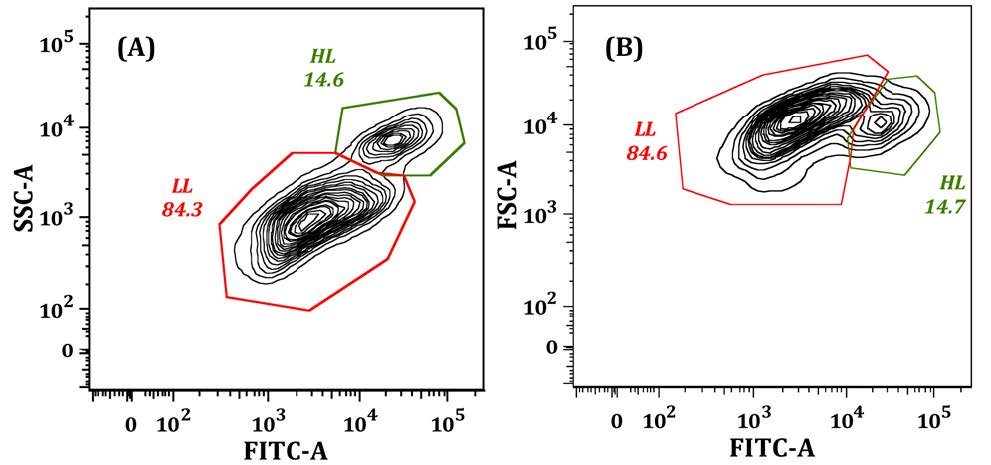
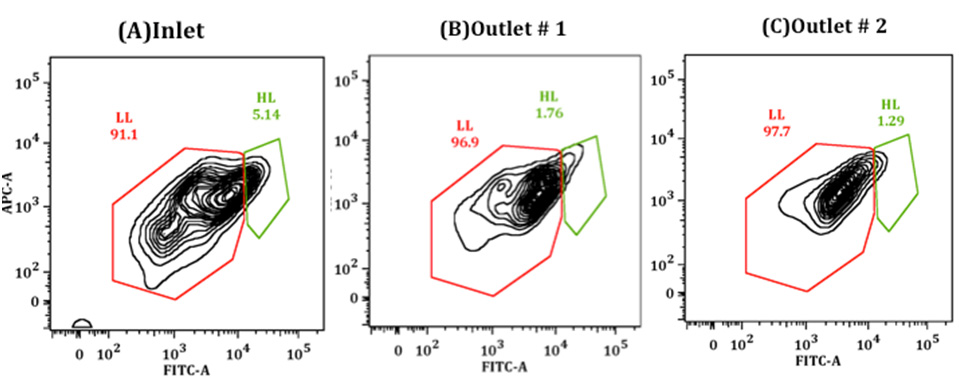
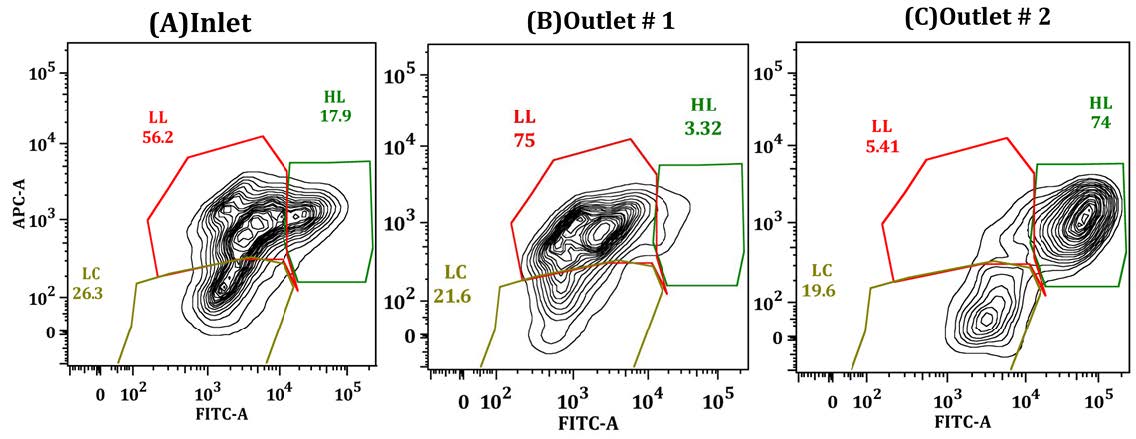
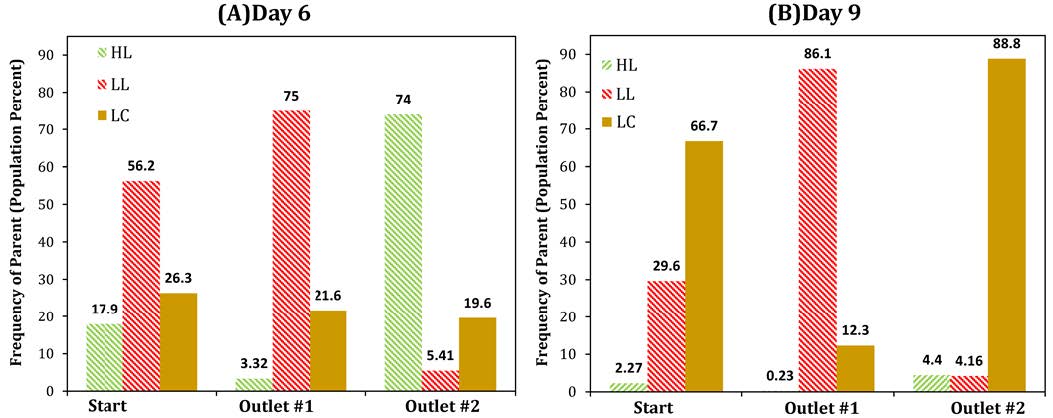
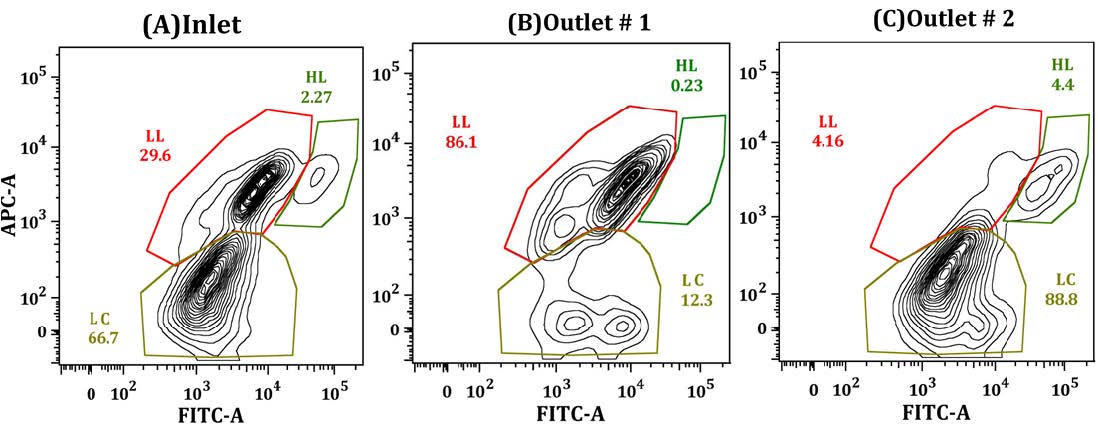
Figures(9)

Hanieh Hadady, Doug Redelman, Sage R. Hiibel, Emil J. Geiger. Continuous-flow sorting of microalgae cells based on lipid content by high frequency dielectrophoresis[J]. AIMS Biophysics, 2016, 3(3): 398-414. doi: 10.3934/biophy.2016.3.398







 DownLoad:
DownLoad: