Citation: Sara R Robertson, Robert JC McLean. Beneficial biofilms[J]. AIMS Bioengineering, 2015, 2(4): 437-448. doi: 10.3934/bioeng.2015.4.437

| [1] |
Vega LM, Alvarez PJ, McLean RJC (2014) Bacterial signaling ecology and potential applications during aquatic biofilm construction. Microbial Ecol 68: 24-34. doi: 10.1007/s00248-013-0321-1

|
| [2] |
Marshall KC, Stout R, Mitchell R (1971) Mechanisms of the initial events in the sorption of marine bacteria to solid surfaces. J Gen Microbiol 68: 337-348. doi: 10.1099/00221287-68-3-337
|
| [3] |
Costerton JW, Geesey GG, Cheng KJ (1978) How bacteria stick. Sci Am 238: 86-95. doi: 10.1038/scientificamerican0178-86
|
| [4] |
Costerton JW, Cheng KJ, Geesey GG, et al. (1987) Bacterial biofilms in nature and disease. Annu Rev Microbiol 41: 435-464. doi: 10.1146/annurev.mi.41.100187.002251
|
| [5] |
Hoiby N (2014) A personal history of research on microbial biofilms and biofilm infections. Pathog Dis 70: 205-211. doi: 10.1111/2049-632X.12165
|
| [6] |
Dobretsov S, Abed RM, Teplitski M (2013) Mini-review: Inhibition of biofouling by marine microorganisms. Biofouling 29: 423-441. doi: 10.1080/08927014.2013.776042
|
| [7] |
Geesey GG, Mutch R, Costerton JW, et al. (1978) Sessile bacteria: an important component of the microbial population in small mountain streams. Limnol Oceanogr 23: 1214-1223. doi: 10.4319/lo.1978.23.6.1214
|
| [8] | Nickel JC, Grant SK, Costerton JW (1985) Catheter associated bacteriuria. An experimental study. Urology 26: 369-375. |
| [9] | Paerl HW, Pinckney JL (1996) A mini-review of microbial consortia: Their roles in aquatic production and biogeochemical cycling. Microb Ecol 31: 225-247. |
| [10] |
Hassanpourfard M, Nikakhtari Z, Ghosh R, et al. (2015) Bacterial floc mediated rapid streamer formation in creeping flows. Sci Rep 5: 13070. doi: 10.1038/srep13070
|
| [11] |
Meyer Reil LA (1994) Microbial life in sedimentary biofilms: The challenge to microbial ecologists. Mar Ecol-Prog Ser 112: 303-311. doi: 10.3354/meps112303
|
| [12] |
Reid RP, Visscher PT, Decho AW, et al. (2000) The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature 406: 989-992. doi: 10.1038/35023158
|
| [13] |
Wentland EJ, Stewart PS, Huang CT, et al. (1996) Spatial variations in growth rate within Klebsiella pneumoniae colonies and biofilm. Biotechnol Prog 12: 316-321. doi: 10.1021/bp9600243
|
| [14] |
Dietrich LE, Okegbe C, Price-Whelan A, et al. (2013) Bacterial community morphogenesis is intimately linked to the intracellular redox state. J Bacteriol 195: 1371-1380. doi: 10.1128/JB.02273-12
|
| [15] | Al-Ahmad A, Wunder A, Auschill TM, et al. (2007) The in vivo dynamics of Streptococcus spp., Actinomyces naeslundii, Fusobacterium nucleatum and Veillonella spp. in dental plaque biofilm as analysed by five-colour multiplex fluorescence in situ hybridization. J Med Microbiol 56: 681-687. |
| [16] | Andersen JB, Sternberg C, Poulsen LK, et al. (1998) New unstable variants of green fluorescent protein for studies of transient gene expression in bacteria. Appl Environ Microbiol 64: 2240-2246. |
| [17] |
Cook LC, Federle MJ (2014) Peptide pheromone signaling in Streptococcus and Enterococcus. FEMS Microbiol Rev 38: 473-492. doi: 10.1111/1574-6976.12046
|
| [18] |
Ng WL, Bassler BL (2009) Bacterial quorum-sensing network architectures. Annu Rev Genet 43: 197-222. doi: 10.1146/annurev-genet-102108-134304
|
| [19] |
Cuadra-Saenz G, Rao DL, Underwood AJ, et al. (2012) Autoinducer-2 influences interactions amongst pioneer colonizing streptococci in oral biofilms. Microbiology 158: 1783-1795. doi: 10.1099/mic.0.057182-0
|
| [20] | Montgomery K, Charlesworth JC, LeBard R, et al. (2013) Quorum sensing in extreme environments. Life (Basel) 3: 131-148. |
| [21] |
Ramage G, Saville SP, Wickes BL, et al. (2002) Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl Environ Microbiol 68: 5459-5463. doi: 10.1128/AEM.68.11.5459-5463.2002
|
| [22] |
Nobbs AH, Jenkinson HF (2015) Interkingdom networking within the oral microbiome. Microbes Infect 17: 484-492. doi: 10.1016/j.micinf.2015.03.008
|
| [23] |
Haruta S, Kato S, Yamamoto K, et al. (2009) Intertwined interspecies relationships: approaches to untangle the microbial network. Environ Microbiol 11: 2963-2969. doi: 10.1111/j.1462-2920.2009.01956.x
|
| [24] | Kouzuma A, Kato S, Watanabe K (2015) Microbial interspecies interactions: recent findings in syntrophic consortia. Front Microbiol 6: 477. |
| [25] |
Schink B (2002) Synergistic interactions in the microbial world. Antonie van Leeuwenhoek 81: 257-261. doi: 10.1023/A:1020579004534
|
| [26] | Juretschko S, Timmermann G, Schmid M, et al. (1998) Combined molecular and conventional analyses of nitrifying bacterium diversity in activated sludge: Nitrosococcus mobilis and Nitrospira- like bacteria as dominant populations. Appl Environ Microbiol 64: 3042-3051. |
| [27] | Boyle M. (1993) Microbial ecology of sewage treatment, In: Ford TE Author, Aquatic microbiology: An ecological approach,. Boston: Blackwell Scientific Publications, 441-454. |
| [28] | Parker DS, Jacobs T, Bower E, et al. (1997) Maximizing trickling filter nitrification rates through biofilm control: Research review and full scale application. Water Sci Technol 36: 255-262. |
| [29] |
Vogelsang C, Husby A, Ostgaard K (1997) Functional stability of temperature-compensated nitrification in domestic wastewater treatment obtained with PVA-SbQ/alginate gel entrapment. Water Res 31: 1659-1664. doi: 10.1016/S0043-1354(97)00009-2
|
| [30] |
Liu H, Logan BE (2004) Electricity generation using an air-cathode single chamber microbial fuel cell in the presence and absence of a proton exchange membrane. Environ Sci Technol 38: 4040-4046. doi: 10.1021/es0499344
|
| [31] |
Rabaey K, Lissens G, Siciliano SD, et al. (2003) A microbial fuel cell capable of converting glucose to electricity at high rate and efficiency. Biotechnol Lett 25: 1531-1535. doi: 10.1023/A:1025484009367
|
| [32] |
Min B, Logan BE (2004) Continuous electricity generation from domestic wastewater and organic substrates in a flat plate microbial fuel cell. Environ Sci Technol 38: 5809-5814. doi: 10.1021/es0491026
|
| [33] | Fangzhou D, Zhenglong L, Shaoqiang Y, et al. (2015) Electricity generation directly using human feces wastewater for life support system. Acta Astronaut 68: 1537-1547. |
| [34] | Gorby YA, Yanina S, McLean JS, et al. (2006) Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other microorganisms. Proc Nat Acad Sci U S A 103: 11358-11363. |
| [35] |
Malvankar NS, Lovley DR (2014) Microbial nanowires for bioenergy applications. Curr Opin Biotechnol 27: 88-95. doi: 10.1016/j.copbio.2013.12.003
|
| [36] |
Wang Y, Kern SE, Newman DK (2010) Endogenous phenazine antibiotics promote anaerobic survival of Pseudomonas aeruginosa via extracellular electron transfer. J Bacteriol 192: 365-369. doi: 10.1128/JB.01188-09
|
| [37] |
Little B, Wagner P (1996) An overview of microbiologically influenced corrosion of metals and alloys. Can J Microbiol 42: 367-374. doi: 10.1139/m96-052
|
| [38] |
Rodriguez-Navarro C, Jroundi F, Schiro M, et al. (2012) Influence of substrate mineralogy on bacterial mineralization of calcium carbonate: implications for stone conservation. Appl Environ Microbiol 78: 4017-4029. doi: 10.1128/AEM.07044-11
|
| [39] |
McLean RJC, Nickel JC, Cheng KJ, et al. (1988) The ecology and pathogenicity of urease-producing bacteria in the urinary tract. Crit Rev Microbiol 16: 37-79. doi: 10.3109/10408418809104467
|
| [40] | Dumanski AJ, Hedelin H, Edin-Liljegren A, et al. (1994) Unique ability of Proteus mirabilis capsule to enhance mineral growth in infectious urinary calculi. Infect Immun 62: 2998-3003. |
| [41] | McLean RJC, Beveridge TJ (1990) Metal binding capacity of bacterial surfaces and their ability to form mineralized aggregates. In: Ehrlich HL, Author, Microbial Mineral Recovery, New York: McGraw-Hill, 185-222. |
| [42] | Egan S, James S, Holmstrom C, et al. (2002) Correlation between pigmentation and antifouling compounds produced by Pseudoalteromonas tunicata. Environ Microbiol: 433-442. |
| [43] |
Maddula VS, Pierson EA, Pierson LS (2008) Altering the ratio of phenazines in Pseudomonas chlororaphis (aureofaciens) strain 30-84: effects on biofilm formation and pathogen inhibition. J Bacteriol 190: 2759-2766. doi: 10.1128/JB.01587-07
|
| [44] |
Human Microbiome Project Consortium (2012) Structure, function and diversity of the healthy human microbiome. Nature 486: 207-214. doi: 10.1038/nature11234
|
| [45] | Yuille S, Mackay WG, Morrison DJ, et al. (2015) Optimising gut colonisation resistance against Clostridium difficile infection. Eur J Clin Microbiol Infect Dis [in press]. |
| [46] |
Cruz MR, Graham CE, Gagliano BC, et al. (2013) Enterococcus faecalis inhibits hyphal morphogenesis and virulence of Candida albicans. Infect Immun 81: 189-200. doi: 10.1128/IAI.00914-12
|
| [47] |
Hogan DA, Kolter R (2002) Pseudomonas-Candida interactions: an ecological role for virulence factors. Science 296: 2229-2232. doi: 10.1126/science.1070784
|
| [48] | Bachtiar EW, Bachtiar BM, Jarosz LM, et al. (2014) AI-2 of Aggregatibacter actinomycetemcomitans inhibits Candida albicans biofilm formation. Front Cell Infect Microbiol 4: 94. |
| [49] | McLean RJC (2014) Normal bacterial flora may inhibit Candida albicans biofilm formation by autoinducer-2. Front Cell Infect Microbiol 4: 117. |
| [50] |
Chung KK, Schumacher JF, Sampson EM, et al. (2007) Impact of engineered surface microtopography on biofilm formation of Staphylococcus aureus. Biointerphases 2: 89-94. doi: 10.1116/1.2751405
|
| [51] | Rogers J, Dowsett AB, Keevil CW (1995) A paint incorporating silver to control mixed biofilms containing Legionella pneumophila. J Indust Microbiol 15: 377-383. |
| [52] | Flemming HC, Griebe T, Schaule G (1996) Antifouling strategies in technical systems: A short review. Water Sci Technol 34: 517-524. |
| [53] |
Breidenstein EB, de la Fuente-Núñez C, Hancock REW (2011) Pseudomonas aeruginosa: all roads lead to resistance. Trends Microbiol 19: 419-426. doi: 10.1016/j.tim.2011.04.005
|
| [54] |
Hentzer M, Riedel K, Rasmussen TB, et al. (2002) Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 148: 87-102. doi: 10.1099/00221287-148-1-87
|
| [55] |
Brackman G, Cos P, Maes L, et al. (2011) Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrob Agents Chemother 55: 2655-2661. doi: 10.1128/AAC.00045-11
|
| [56] |
Davies DG, Marques CNH (2009) A fatty acid messenger is responsible for inducing dispersion in microbial biofilms. J Bacteriol 191: 1393-1403. doi: 10.1128/JB.01214-08
|
| [57] | Lanter BB, Sauer K, Davies DG (2014) Bacteria present in carotid arterial plaques are found as biofilm deposits which may contribute to enhanced risk of plaque rupture. mBio 5: e01206-14. |
| [58] |
Nicolaou SA, Gaida SM, Papoutsakis ET (2010) A comparative view of metabolite and substrate stress and tolerance in microbial bioprocessing: From biofuels and chemicals, to biocatalysis and bioremediation. Metab Eng 12: 307-331. doi: 10.1016/j.ymben.2010.03.004
|
| [59] | Beijerinck MW. (1999) Enrichment culture studies with urea bacteria (translation of original article “Anhäufungsversuche mit ureumbakterien.” Centralblatt für Bakteriologie, Part II, Vol 7, pp 33-61, 1901) In: Milestones in microbiology: 1546 to 1940, Washington DC: ASM Press, 234-237. |
| [60] | Kolenbrander PE, London J (1993) Adhere today, here tomorrow: Oral bacterial adherence. J Bacteriol 175: 3247-3252. |
| [61] |
Jang YJ, Sim J, Jun HK, et al. (2013) Differential effect of autoinducer 2 of Fusobacterium nucleatum on oral streptococci. Arch Oral Biol 58: 1594-1602. doi: 10.1016/j.archoralbio.2013.08.006
|
| [62] |
Ferrières L, Hancock V, Klemm P (2007) Specific selection for virulent urinary tract infectious Escherichia coli strains during catheter-associated biofilm formation. FEMS Immunol Med Microbiol 51: 212-219. doi: 10.1111/j.1574-695X.2007.00296.x
|
| [63] |
Ishida KP, Griffiths PR (1999) Investigation of polysaccharide adsorption on protein conditioning films by attenuated total reflection infrared spectrometry - II. Thin copper films. J Colloid Interface Sci 213: 513-524. doi: 10.1006/jcis.1999.6162
|
| [64] |
Zhang N, Yang D, Wang D, et al. (2015) Whole transcriptomic analysis of the plant-beneficial rhizobacterium Bacillus amyloliquefaciens SQR9 during enhanced biofilm formation regulated by maize root exudates. BMC Genomics 16: 685. doi: 10.1186/s12864-015-1825-5
|
| [65] |
Chaparro JM, Badri DV, Bakker MG, et al. (2013) Root exudation of phytochemicals in Arabidopsis follows specific patterns that are developmentally programmed and correlate with soil microbial functions. PLOS One 8: e55731. doi: 10.1371/journal.pone.0055731
|
| [66] |
Egan S, Harder T, Burke C, et al. (2013) The seaweed holobiont: understanding seaweed-bacteria interactions. FEMS Microbiol Rev 37: 462-476. doi: 10.1111/1574-6976.12011
|
| [67] | Givskov M, de Nys R, Manefield M, et al. (1996) Eukaryotic interference with homoserine lactone-mediated prokaryotic signalling. J Bacteriol 178: 6618-6622. |
| [68] |
McLean RJC, Barnes MB, Windham MK, et al. (2005) Cell-cell influences on bacterial community development in aquatic biofilms. Appl Environ Microbiol 71: 8987-8990. doi: 10.1128/AEM.71.12.8987-8990.2005
|
| [69] |
Nickel JC, Ruseska I, Wright JB, et al. (1985) Tobramycin resistance of Pseudomonas aeruginosa cells growing as a biofilm on urinary catheter material. Antimicrob Agents Chemother 27: 619-624. doi: 10.1128/AAC.27.4.619
|
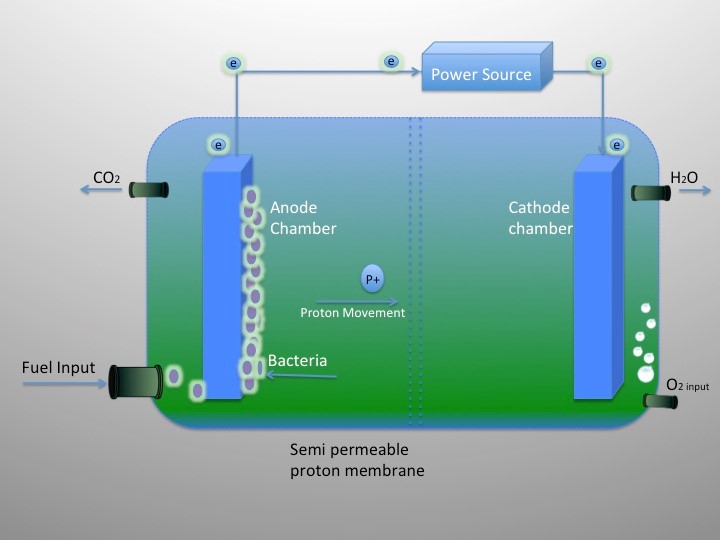
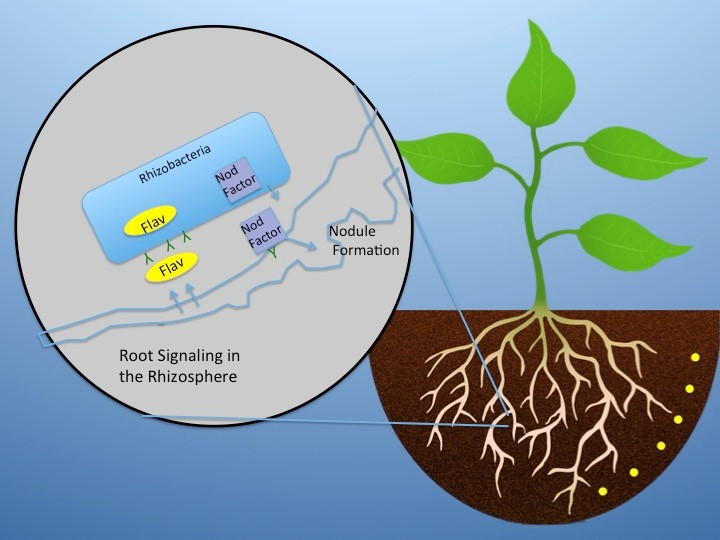
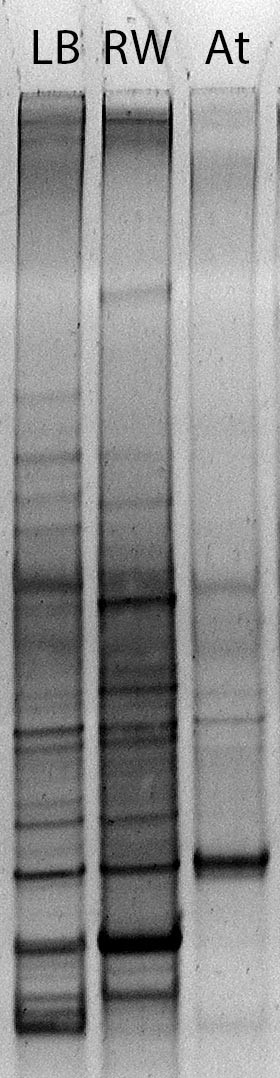
Figures(3)

Sara R Robertson, Robert JC McLean. Beneficial biofilms[J]. AIMS Bioengineering, 2015, 2(4): 437-448. doi: 10.3934/bioeng.2015.4.437







 DownLoad:
DownLoad: