Citation: Yaguang Huangfu, Guanqing Liang, Jiannong Cao. MatrixMap: Programming abstraction and implementation of matrix computation for big data analytics[J]. Big Data and Information Analytics, 2016, 1(4): 349-376. doi: 10.3934/bdia.2016015

| [1] | [ C.-C. Chang and Chih-Jen, libsvm dataset url:http://www.csie.ntu.edu.tw/~cjlin/libsvmtools/datasets/binary/news20.binary.bz2, 2015. |
| [2] | [ J. Choi, J. J. Dongarra, R. Pozo and D. W. Walker, ScaLAPACK:A scalable linear algebra library for distributed memory concurrent computers, in Frontiers of Massively Parallel Computation, 1992., Fourth Symposium on the, IEEE, (1992), 120-127. |
| [3] | [ Chu, Cheng-Tao and Kim, Sang Kyun and Lin, Yi-An and Yu, YuanYuan and Bradski, Gary and Ng, Andrew Y and Olukotun, Kunle, Map-Reduce for Machine Learning on Multicore, in Neural Information Processing Systems, 2007. |
| [4] | [ M. T. Chu and J. L. Watterson, On a multivariate eigenvalue problem, Part I:Algebraic theory and a power method, SIAM Journal on Scientific Computing, 14(1993), 1089-1106. |
| [5] | [ T. H. Cormen, Introduction to Algorithms, MIT press, 2009. |
| [6] | [ J. Dean and S. Ghemawat, MapReduce:simplified data processing on large clusters, Communications of the ACM, 51(2008), 107-113. |
| [7] | [ J. Ekanayake, H. Li and B. Zhang, Twister:A runtime for iterative MapReduce, in HPDC'10 Proceedings of the 19th ACM International Symposium on High Performance Distributed Computing, (2010), 810-818. |
| [8] | [ J. Gonzalez, Y. Low, H. Gu, D. Bickson and C. Guestrin, PowerGraph:Distributed graphparallel computation on natural graphs, in OSDI'12 Proceedings of the 10th USENIX conference on Operating Systems Design and Implementation, (2012), 17-30. |
| [9] | [ P. Harrington, Machine Learning in Action, Manning Publications, 2012. |
| [10] | [ P. Hintjens, ZeroMQ:Messaging for Many Applications, O'Reilly Media, Inc., 2013. |
| [11] | [ Intel, Threading Building Blocks url:https://www.threadingbuildingblocks.org/, 2009. |
| [12] | [ M. Isard, M. Budiu, Y. Yu, A. Birrell and D. Fetterly, Dryad:distributed data-parallel programs from sequential building blocks, ACM SIGOPS Operating Systems Review, 41(2007), 59-72. |
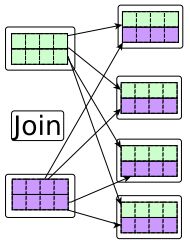
| [13] | [ Join (SQL) url:https://en.wikipedia.org/wiki/Join, 2015. |
| [14] | [ J. Kepner and J. Gilbert, Graph Algorithms in the Language of Linear Algebra, SIAM, 2011. |
| [15] | [ K. Kourtis, V. Karakasis, G. Goumas and N. Koziris, CSX:An extended compression format for spmv on shared memory systems, in ACM SIGPLAN Notices, 46(2011), 247-256. |
| [16] | [ J. Kowalik, ACTORS:A model of concurrent computation in distributed systems (Gul Agha), SIAM Review, 30(1988), 146-146. |
| [17] | [ C. G. Aapo Kyrola and G. Blelloch, GraphChi:Large-scale graph computation on just a PC, in Proceedings of the 10th USENIX conference on Operating Systems Design and Implementation, USENIX Association, (2012), 31-46. |
| [18] | [ Y. Low, J. Gonzalez and A. Kyrola, Graphlab:A distributed framework for machine learning in the cloud, arXiv preprint, arXiv:1107.0922, 1107(2011). |
| [19] | [ Y. Low, J. Gonzalez, A. Kyrola, D. Bickson, C. Guestrin and J. M. Hellerstein, Distributed GraphLab:A Framework for Machine Learning and Data Mining in the Cloud, in Proceedings of the VLDB Endowment, 5(2012), 716-727. |
| [20] | [ G. Malewicz, M. Austern and A. Bik, Pregel:A system for large-scale graph processing, Proceedings of the the 2010 international conference on Management of data, 114(2010), 135-145. |
| [21] | [ D. Murray, F. McSherry, R. Isaacs, M. Isard, P. Barham and M. Abadi, Naiad:A timely dataflow system, in SOSP'13:Proceedings of the Twenty-Fourth ACM Symposium on Operating Systems Principles, (2013), 439-455. |
| [22] | [ E. J. O'Neil, P. E. O'Neil and G. Weikum, The LRU-K page replacement algorithm for database disk buffering, in ACM SIGMOD Record, 22(1993), 297-306. |
| [23] | [ T. W. L Page, S Brin, R Motwani, The PageRank Citation Ranking:Bringing Order to the Web, tech. rep., Stanford InfoLab, 1999. |
| [24] | [ R. Power and J. Li, Piccolo:Building fast, distributed programs with partitioned tables, Proceedings of the 9th USENIX conference on Operating systems design and implementation-OSDI'10, (2010), 1-14. |
| [25] | [ J. Protic, M. Tomasevic and V. Milutinovi´c, Distributed Shared Memory:Concepts and Systems, John Wiley & Sons, 1998. |
| [26] | [ Z. Qian, X. Chen, N. Kang and M. Chen, MadLINQ:large-scale distributed matrix computation for the cloud, Proceedings of the 7th ACM european conference on Computer Systems. ACM, (2012), 197-210,. |
| [27] | [ RocksDB, http://rocksdb.org/, 2015. |
| [28] | [ A. Roy, I. Mihailovic and W. Zwaenepoel, X-stream:edge-centric graph processing using streaming partitions, in the Twenty-Fourth ACM Symposium on Operating Systems Principles, (2013), 472-488. |
| [29] | [ S. Seo, E. J. Yoon, J. Kim, S. Jin, J.-S. Kim and S. Maeng, HAMA:An efficient matrix computation with the mapreduce framework, in 2010 IEEE Second International Conference on Cloud Computing Technology and Science, (2010), 721-726. |
| [30] | [ J. Shun and G. Blelloch, Ligra:A lightweight graph processing framework for shared memory, in PPoPP, (2013), 135-146. |
| [31] | [ M. S. Snir, S. W. Otto, D. W. Walker, J. Dongarra and Huss-Lederman, MPI:The Complete Reference, MIT Press, 1995. |
| [32] | [ L. Valiant, A bridging model for parallel computation, Communications of the ACM, 33(1990), 103-111. |
| [33] | [ P. Vassiliadis, A survey of extract-transform-load technology, International Journal of Data Warehousing and Mining, 5, 1-27. |
| [34] | [ S. Venkataraman, E. Bodzsar, I. Roy, A. AuYoung, and R. S. Schreiber, Presto, in Proceedings of the 8th ACM European Conference on Computer Systems-EuroSys'13, (2013), p197. |
| [35] | [ R. S. Xin, J. E. Gonzalez, M. J. Franklin, I. Stoica, and E. AMPLab, GraphX:A Resilient Distributed Graph System on Spark, in First International Workshop on Graph Data Management Experiences and Systems, p. 2, 2013. |
| [36] | [ M. Zaharia, M. Chowdhury, M. J. Franklin, S. Shenker and I. Stoica, Spark:Cluster computing with working sets, HotCloud'10 Proceedings of the 2nd USENIX conference on Hot topics in cloud computing, (2010), p10. |
| [37] | [ M. Zaharia, M. Chowdhury, T. Das and A. Dave, Resilient Distributed Datasets:A FaultTolerant Abstraction for In-Memory Cluster Computing, tech. rep., UCB/EECS-2011-82 UC Berkerly, 2012. |
| [38] | [ T. Zhang, Solving large scale linear prediction problems using stochastic gradient descent algorithms, in Proceedings of the twenty-first international conference on Machine learning, ACM, (2004), p116. |
| [39] | [ Y. Zhou, D. Wilkinson, R. Schreiber and R. Pan, Large-scale parallel collaborative filtering for the netflix prize, Lecture Notes in Computer Science (including subseries Lecture Notes in Artificial Intelligence and Lecture Notes in Bioinformatics), LNCS, 5034(2008), 337-348. |
Figures(13) / Tables(10)

Yaguang Huangfu, Guanqing Liang, Jiannong Cao. MatrixMap: Programming abstraction and implementation of matrix computation for big data analytics[J]. Big Data and Information Analytics, 2016, 1(4): 349-376. doi: 10.3934/bdia.2016015









 DownLoad:
DownLoad: