Citation: Enrica Bargiacchi, Margherita Campo, Annalisa Romani, Gilberto Milli, Sergio Miele. Hydrolysable Tannins from Sweet Chestnut (Castanea sativa Mill.) to improve Tobacco and Food/Feed Quality. Note 1: Fraction characterization, and Tobacco biostimulant effect for gall-nematode resistance[J]. AIMS Agriculture and Food, 2017, 2(3): 324-338. doi: 10.3934/agrfood.2017.3.324

| [1] | Lorenzo JM, González-Rodríguez RM, Sánchez M, et al. (2013) Effects of natural (grape seed and chestnut extract) and synthetic antioxidants (buthylatedhydroxytoluene, BHT) on the physical, chemical. Microbiological and sensory characteristics of dry cured sausage "chorizo". Food Res Int 54: 611-620. |
| [2] | Pateiro M, Lorenzo JM, Amado IR, et al. (2014) Effect of addition of green tea, chestnut and grape extract on the shelf-life of pig liver pâté. Food Chem 147: 386-394. |
| [3] |
Liu HW, Dong XF, Tong JM, et al. (2011) A comparative study of growth performance and antioxidant status of rabbits when fed with or without chestnut tannins under high ambient temperature. Anim Feed Sci Tech 164: 89-95. doi: 10.1016/j.anifeedsci.2010.09.020

|
| [4] | Frankič T, Salobir J (2011) In vivo antioxidant potential of Sweet chestnut (Castanea sativa Mill.) wood extract in young growing pigs exposed to n-3 PUFA-induced oxidative stress. J Sci Food Agr 91:1432-1439. |
| [5] | Živković J, Zeković Z, Mujić I, et al. (2010) Scavenging capacity of superoxide radical and screening of antimicrobial activity of Castanea sativa Mill. extracts. Czech J Food Sci 1: 61-68. |
| [6] |
Buzzini P, Arapitsas P, Goretti M, et al. (2008) Antimicrobial and antiviral activity of hydrolysable tannins. Mini-Rev Med Chem 8: 1179-1187. doi: 10.2174/138955708786140990
|
| [7] | Romani A, Campo M, Pinelli P, et al. (2010) Hydrolyzable tannins from different vegetal species, fractionation HPLC/DAD/MS analyses, and anti-yeast activity. XXVth international Conference on Polyphenols, Polyphenols Communication 2010, Vol. II, 611-612. |
| [8] |
Elizondo AM, Mercado EC, Rabinovitz BC, et al. (2010) Effect of tannins on the in vitro growth of Clostridium perfringens. Vet Microbiol 145: 308-314. doi: 10.1016/j.vetmic.2010.04.003
|
| [9] |
Budriesi R, Ioan P, Micucci M, et al. (2010) Stop Fitan: Antispasmodic effect of natural extract of Chestnut wood in Guinea pig ileum and proximal colon smooth muscle. J Med Food 13: 1104-1110. doi: 10.1089/jmf.2009.0210
|
| [10] | Bargiacchi E, Bertola R, Costa G, et al. (2010) Method for obtaining organic acidifying solutions and fertilizers using organic extracts from aqueous wood leaching processes. Eur. Pat. EP1464635, 24-03-2010, priority: IT2003MI00640 2003-03-31. |
| [11] |
Martinez S, Štagljar I (2003) Correlation between the molecular structure and the corrosion inhibition efficiency of chestnut tannin in acidic solution. J Mol Struct: THEOCHEM 640: 167-174. doi: 10.1016/j.theochem.2003.08.126
|
| [12] | Rahim AA, Kassim J (2008) Recent development of vegetal tannins in corrosion protection of iron and steel. Recent Patents on Mater Sci 1: 223-231. |
| [13] | Andrade RG; Dalvi LT, Silva JMC, et al. (2005) The antioxidant effect of tannic acid on the in vitro copper mediated formation of free radicals. Arch Biochem Biophys 437: 1-9. |
| [14] | Bargiacchi E, Miele S, Romani A, et al. (2013) Biostimulant activity of hydrolyzable tannins from Sweet Chestnut (Castanea sativa Mill.). Acta Hortic 1009: 111-115. |
| [15] | Maistrello L, Vaccari G, Sasanelli N (2010) Effect of Chestnut tannins on the root-knot nematode Meloidogyne javanica. Helminthologia 47: 48-57. |
| [16] | Rencŏ M, Sasanelli N, Papajová I, et al. (2012) Nematicidal effect of chestnut tannin solutions on the potato cyst nematode Globodera rostochiensis (Woll.) Barhens. Helminthologia 49: 108-114. |
| [17] | Rencŏ M, Sasanelli N, Maistrello L (2014) Plants as natural sources of nematicides, In: Lee M, Davis, Authors, Plants as natural sources of nematicides, New York: NOVA Science publisher, 115-142. |
| [18] | Faizi S, Fayyaz S, Bano S, et al. (2011) Isolation of Nematicidal Compounds from Tagetes patula L. Yellow Flowers: Structure_Activity Relationship Studies against Cyst Nematode Heterodera zeae Infective Stage Larvae. J Agr Food Chem 59: 9080-9093. |
| [19] | Romani A, Campo M, Pinelli P (2012) HPLC/DAD/ESI-MS analyses and anti-radical activity of hydrolyzable tannins from different vegetal species. Food Chem 30: 214-221. |
| [20] | Regione Umbria. Allegato A. Disciplinari di produzione integrate. Sezione Pratiche Agronomiche. Available from: www.agricoltura.regione.umbria.it/mediacenter/ |
| [21] | Barker KR (1985) Nematode extraction and bioassays, In: K R Barker, J N. Sasser, C C Carter, An advanced treatise on Meloidogyne, North Carolina State University Graphics, Raleigh, NC, 19-35. |
| [22] | Campo M, Pinelli P, Romani A (2016) Hydrolyzable Tannins from Sweet Chestnut Fractions Obtained by a Sustainable and Eco-friendly Industrial Process. Nat Prod Commun 11: 409-415. |
| [23] | Sanz M, Cadahìa E, Esteruelas E, et al. (2010) Phenolic compounds in Chestnut (Castanea sativa Mill.) heartwood. Effect on toasting at cooperage. J Agr Food Chem 58: 9631-9640. |
| [24] | Romani A, Campo M, Pinelli P (2012) HPLC/DAD/ESI-MS analyses and anti-radical activity of hydrolyzable tannins from different vegetal species. Food Chem 30: 214-221. |
| [25] | Aaby K, Ekeberg D, Skrede G (2007) Characterization of phenolic compounds in strawberry (Fragaria x ananassa) fruits by different HPLC detectors and contribution of individual compounds to total antioxidant capacity. J Agr Food Chem 55: 4395-4406. |
| [26] | Haddock EA, Gupta RK, Al Shafi SMK, et al. (1982) The metabolism of gallic acid and hexahydroxydiphenic acid in plants: biogenic and molecular taxonomic considerations. Phytochemistry 21: 1049-1062. |
| [27] | Fukuda T, Ito H, Yoshida T (2003) Antioxidative polyphenols from walnuts (Juglans regia L.). Phytochemistry 63: 795-801. |
| [28] | Comandini P, Lerma-García MJ, Simó-Alfonso EF, et al. (2014) Tannin analysis of chestnut bark samples (Castanea sativa Mill.) by HPLC-DAD-MS. Food Chem 157: 290-295. |
| [29] | Cantos E, Espìn JC, Lòpez-Bote C, et al. (2003) Phenolic compounds and fatty acids from Acorns (Quercus spp.), the main dietary constituent of free-ranged Iberian pigs. J Agr Food Chem 51: 6248-6255. |
| [30] | Fernandes A, Sousa A, Mateus N, et al. (2011) Analysis of cork from Quercus suber L. by HPLC-DAD/ESI-MS. Food Chem 125: 1398-1405. |
| [31] | Quéméner B, Ralet MC (2004) Evidence for linkage position determination in known feruloylated mono- and disaccharides using electrospray ion trap mass spectrometry. J Mass Spectrome 39: 1153-1160. |
| [32] | Meyers KJ, Tedmund JS, Mitchell AE (2006) Understanding the native Californian diet: identification of condensed and hydrolysable tannins in Tanoak acorns (Lithocarpus densiflorus). J Agr Food Chem 54: 7686-7691. |
| [33] | Arapitsas P, Menichetti S, Vincieri FF, et al. (2007) Hydrolyzable Tannins with the Hexahydroxydiphenoyl unit and the m-depsidic link: HPLC-DAD-MS identification and model synthesis. J Agr Food Chem 55: 48-55. |
| [34] | Garcia Izquierdo C, 2016. Evergreen B6, C2 and C6 Deliverables. Life2013 ENV IT 000461/ "Evergreen" personal comm. |
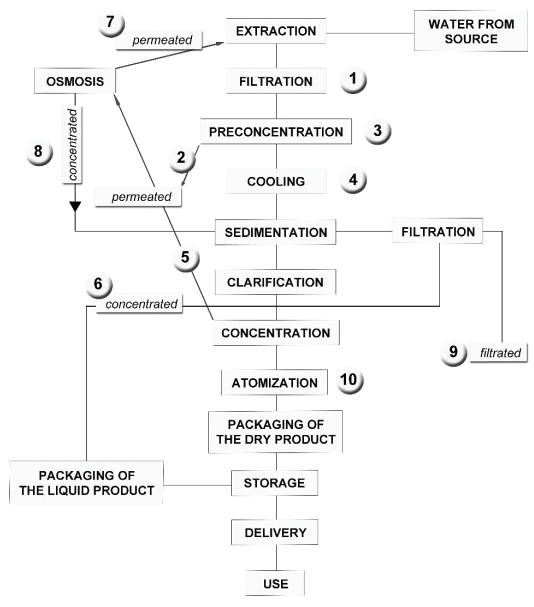
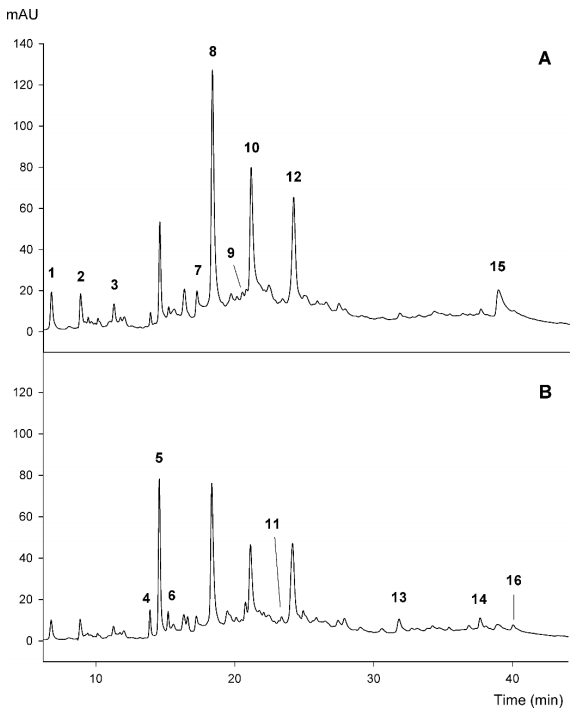
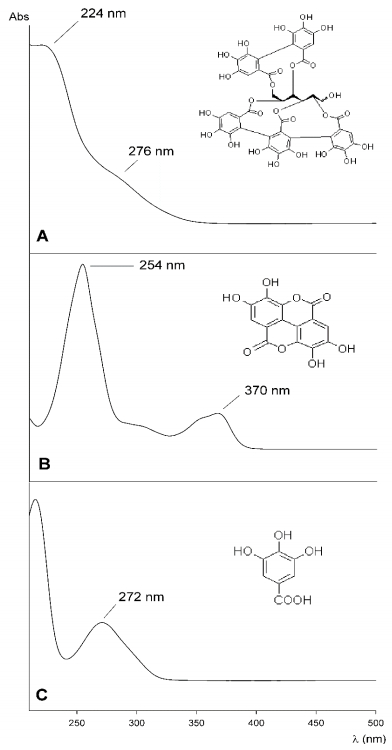
Figures(3) / Tables(6)

Enrica Bargiacchi, Margherita Campo, Annalisa Romani, Gilberto Milli, Sergio Miele. Hydrolysable Tannins from Sweet Chestnut (Castanea sativa Mill.) to improve Tobacco and Food/Feed Quality. Note 1: Fraction characterization, and Tobacco biostimulant effect for gall-nematode resistance[J]. AIMS Agriculture and Food, 2017, 2(3): 324-338. doi: 10.3934/agrfood.2017.3.324







 DownLoad:
DownLoad: