Citation: Cíntia Sorane Good Kitzberger, Maria Brígida dos Santos Scholz, Luiz Filipe Protasio Pereira, João Batista Gonçalves Dias da Silva, Marta de Toledo Benassi. Profile of the diterpenes, lipid and protein content of different coffee cultivars of three consecutive harvests[J]. AIMS Agriculture and Food, 2016, 1(3): 254-264. doi: 10.3934/agrfood.2016.3.254

| [1] | Navarini L, Ferrari M, Liverani FS, et al. (2004) Dynamic tensiometric characterization of espresso coffee beverage. Food Hydrocoll 18: 387-393. |
| [2] | Piazza L, Bulbarello A, Gigli J (2006) Rheological interfacial properties of espresso coffee foaming fractions. In Proceedings of the IUFoST 13th World Congress of Food Science & Technology, France. |
| [3] |
Nebesny E, Budryn G (2006) Evaluation of sensory attributes of coffee brews from robusta coffee roasted under different conditions. Eur Food Res Technol 224: 159-165. doi: 10.1007/s00217-006-0308-y

|
| [4] |
Farah A, Monteiro MC, Calado V, et al. (2006) Correlation between cup quality and chemical attributes of Brazilian coffee. Food Chem 98: 373-380. doi: 10.1016/j.foodchem.2005.07.032
|
| [5] |
Ramalakshmi K, Kubra IR, Rao LJM (2007) Physicochemical characteristics of green coffee: Comparison of graded and defective beans. J Food Sci 72: S333-S337. doi: 10.1111/j.1750-3841.2007.00379.x
|
| [6] | Scholz MBS, Figueiredo VRG, Silva JVN, et al. (2011) Características físico-químicas de grãos verdes e torrados de cultivares de café (Coffea arabica L.) do Iapar. Coffee Sci 6: 245-255. |
| [7] |
Joet T, Laffarge A, Descroix F, et al. (2010) Influence of environmental factors, wet processing and their interactions on the biochemical composition of green coffee beans. Food Chem 118: 693-701. doi: 10.1016/j.foodchem.2009.05.048
|
| [8] | Speer K, Kölling-Speer I (2006) The lipid fraction of the coffee bean – Minireview. Braz J Plant Physiol 18: 201-216. |
| [9] |
Muriel P, Arauz J (2010) Coffee and liver diseases. Fitoterapia 81: 297-305. doi: 10.1016/j.fitote.2009.10.003
|
| [10] |
Chu YF, Chen Y, Black RM, et al. (2011) Type 2 diabetes-related bioactivities of coffee: Assessment of antioxidant activity, NF-kB inhibition, and stimulation of glucose uptake. Food Chem 124: 914-920. doi: 10.1016/j.foodchem.2010.07.019
|
| [11] |
Lee KJ, Choi JH, Jeong HG (2007) Hepatoprotective and antioxidant effects of the coffee diterpenes kahweol and cafestol on carbon tetrachloride-induced liver damage in mice. Food Chem Toxicol 45: 2118-2125. doi: 10.1016/j.fct.2007.05.010
|
| [12] | Sridevi V, Giridhar P, Ravishankar GA (2010) Free diterpenes cafestol and kahweol in beans and in vitro cultures of Coffea species. Curr Sci 99: 1101-1104. |
| [13] | Sridevi V, Giridhar P (2015) Variations in diterpenes-cafestol and kahweol content in beans of robusta coffee grown at different altitudes. In Proceedings of the National Academy of Sciences, Section B: Biological Sciences, India. |
| [14] |
Cruchten STJ, Van De Haanc LHJ, Mulderd PPJ, et al. (2010) The role of epoxidation and electrophile-responsive element-regulated gene transcription in the potentially beneficial and harmful effects of the coffee components cafestol and kahweol. J Nutr Biochem 21: 757-763. doi: 10.1016/j.jnutbio.2009.05.001
|
| [15] | Ky CL, Louarn J, Dussert S, et al. (2001) Caffeine, trigonelline, chlorogenic acid and sucrose diversity in wild Coffea arabica L. and C. canephora P. accessions. Food Chem 75: 223-230. |
| [16] |
Silva EA, Mazzafera P, Brunini O, et al. (2005) The influence of water management and environmental conditions on the chemical composition and beverage quality of coffee beans. Braz J Plant Physiol 17: 229-238. doi: 10.1590/S1677-04202005000200006
|
| [17] |
Leroy T, Ribeyre F, Bertrand B, et al. (2006) Genetics of coffee quality. Braz J Plant Physiol 18: 229-242. doi: 10.1590/S1677-04202006000100016
|
| [18] | Läderach P, Collet L, Oberthür T, et al. (2006) Relationships between environmental factors and coffee cup quality in selected growing regions of Cauca, southern Colombia. In: 21 International Conference of Coffee Science, Montpellier. 1-12. |
| [19] |
Roos B, Van Der Weg G, Urgert R, et al. (1997) Levels of cafestol, kahweol, and related diterpenoids in wild species of the coffee plant Coffea. J Agric Food Chem 45: 3065-3069. doi: 10.1021/jf9700900
|
| [20] |
Kitzberger CSG, Scholz MBS, Pereira LFP, et al. (2013) Diterpenes in green and roasted coffee of Coffea arabica cultivars growing in the same edapho-climatic conditions. J Food Comp Anal 30: 52-57. doi: 10.1016/j.jfca.2013.01.007
|
| [21] |
Kitzberger CSG, Scholz MBS, Pereira LFP, et al. (2013) Composição química de cafés árabica de cultivares tradicionais e modernas. Pesqui Agropecu Bras 48: 1498-1506. doi: 10.1590/S0100-204X2013001100011
|
| [22] |
Scholz MBS, Pagiatto NF, Kitzberger CSG, et al. (2014) Validation of near-infrared spectroscopy for the quantification of cafestol and kahweol in green coffee. Food Res Int 61: 176-182. doi: 10.1016/j.foodres.2013.12.008
|
| [23] |
Rogers WJ, Bézard G, Deshayes A, et al. (1999) Biochemical and molecular characterization and expression of the 11s-type storage protein from Coffea arabica endosperm. Plant Physiol Biochem 37: 261-272. doi: 10.1016/S0981-9428(99)80024-2
|
| [24] |
Montavón P, Mauron AF, Duruz E (2003) Changes in green coffee protein profiles during roasting. J Agric Food Chem 51: 2335-2343. doi: 10.1021/jf020832b
|
| [25] | Ali MKA (2013) Interactions of food proteins with plant phenolics – modulation of structural, techno- and bio-functional properties of proteins. PhD Thesis, Wissenschaftsdiziplin Ernährungswissenschaft eingereicht von Betreuer, Potsdam. |
| [26] | Rodrigues CI, Maia R, Máguas C (2010) Comparing total nitrogen and crude protein content of green coffee beans (Coffea spp.) from different geographical origins. Coffee Sci 5: 197-205. |
| [27] |
Hernández JA, Heyd B, Irles C, et al. (2007) Analysis of the heat and mass transfer during coffee batch roasting. J Food Eng 78: 1141-1148. doi: 10.1016/j.jfoodeng.2005.12.041
|
| [28] |
De Maria CAB, Trugo LC, Neto FRA, et al. (1996) Composition of green coffee water-soluble fractions and identification of volatiles formed during roasting. Food Chem 55: 203-207. doi: 10.1016/0308-8146(95)00104-2
|
| [29] |
Bekedam EK, Schols HA, Van Boekel MAJS, et al. (2006) High molecular weight melanoidins from coffee brew. J Agric Food Chem 54: 7658-7666. doi: 10.1021/jf0615449
|
| [30] |
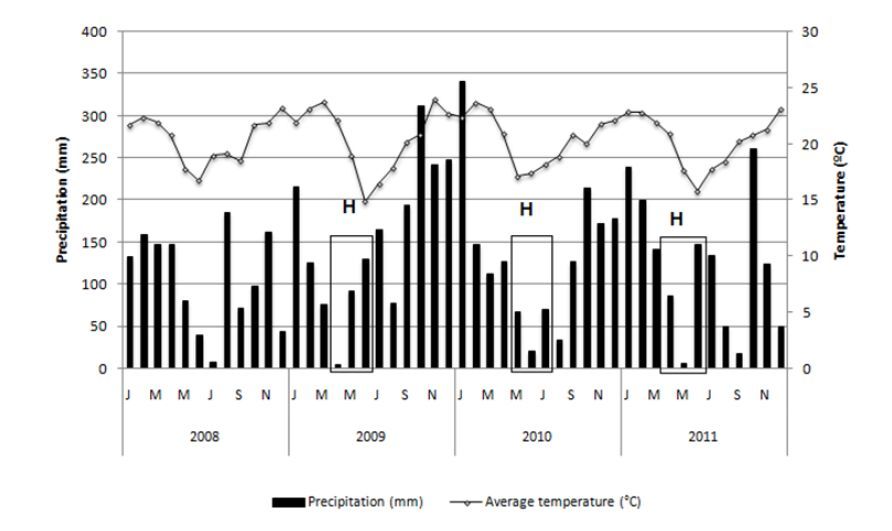
Pezzopane JR, Salva TJG, Lima VB, et al. (2012) Agrometeorological parameters for prediction of the maturation period of Arabica coffee cultivars. Int J Biometeorol 56: 843-851. doi: 10.1007/s00484-011-0486-6
|
| [31] |
Ito DS, Sera T, Sera GH, et al. (2008) Resistance to bacterial blight in arabica coffee cultivars. Crop Breed Appl Biot 8: 99-103. doi: 10.12702/1984-7033.v08n02a01
|
| [32] | Caramori PH, Caviglione JH, Wrege MS, et al. (2001) Zoneamento de riscos climáticos para a cultura de café (Coffea arabica L.) no estado do Paraná. Rev Bras Meteorol 9: 486-494. |
| [33] | BRASIL (2003) Ministério da Agricultura, Pecuária e Abastecimento. Regulamento técnico de identidade e de qualidade para a classificação do café beneficiado e do grão verde. Instrução Normativa n°8 de 11 de jun. 2003. |
| [34] |
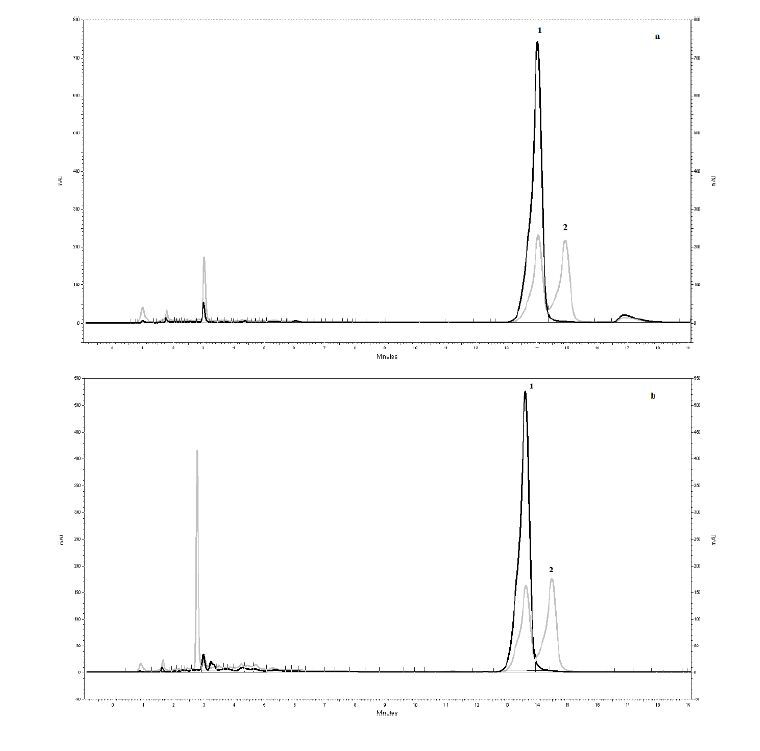
Dias RCE, Campanha FG, Vieira LGE, et al. (2010) Evaluation of kahweol and cafestol in coffee tissues and roasted coffee by a new high-performance liquid chromatography methodology. J Agric Food Chem 58: 88-93. doi: 10.1021/jf9027427
|
| [35] | Association of Official Analytical Chemists (1990) Official methods of analysis. 15th ed. Arlington. 1298. |
| [36] | Scholz MBS, Kitzberger CSG, Pereira LFP, et al. (2014) Application of near infrared spectroscopy for green coffee biochemical phenotyping. J Near Infrared Spectrosc 22: 411-421. |
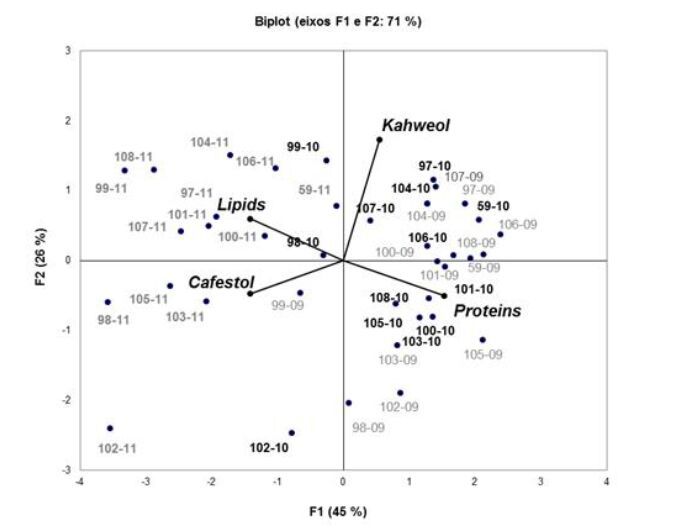
| [37] | Addinsoft (2010) XLStat: software for statistical analysis. Versião 2008.4.02 (2008). Paris. 1 CD-ROM |
| [38] |
Geromel C, Ferreira LP, Davrieux F, et al. (2008) Effects of shade on the development and sugar metabolism of coffee (Coffea arabica L.) fruits. Plant Physiol Biochem 46: 569-579. doi: 10.1016/j.plaphy.2008.02.006
|
| [39] | Mazzafera P, Soave D, Zullo MAT, et al. (1998) Oil content of green coffee beans from some coffee species. Bragantia 57: 45-48. |
| [40] |
Oliveira LS, Franca AS, Mendonça JCF, et al. (2006) Proximate composition and fatty acids profile of green and roasted defective coffee beans. LWT Food Sci Technol 39: 235-239. doi: 10.1016/j.lwt.2005.01.011
|
| [41] |
Butt MS, Sultan MT (2011) Coffee and its consumption: benefits and risks. Crit Rev Food Sci Nutr 51: 363-373. doi: 10.1080/10408390903586412
|
Figures(3) / Tables(4)

Cíntia Sorane Good Kitzberger, Maria Brígida dos Santos Scholz, Luiz Filipe Protasio Pereira, João Batista Gonçalves Dias da Silva, Marta de Toledo Benassi. Profile of the diterpenes, lipid and protein content of different coffee cultivars of three consecutive harvests[J]. AIMS Agriculture and Food, 2016, 1(3): 254-264. doi: 10.3934/agrfood.2016.3.254







 DownLoad:
DownLoad: