Citation: Ellie Sayers, Alex MacGregor, Simon R. Carding. Drug-microbiota interactions and treatment response: Relevance to rheumatoid arthritis[J]. AIMS Microbiology, 2018, 4(4): 642-654. doi: 10.3934/microbiol.2018.4.642

| [1] | Carding S, Verbeke K, Vipond DT, et al. (2015) Dysbiosis of the gut microbiota in disease. Microb Ecol Heal Dis 26: 26191–26199. |
| [2] |
Scher JU, Sczesnak A, Longman RS, et al. (2013) Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. Elife 2: e01202. doi: 10.7554/eLife.01202

|
| [3] |
Liu X, Zou Q, Zeng B, et al. (2013) Analysis of fecal Lactobacillus community structure in patients with early rheumatoid arthritis. Curr Microbiol 67: 170–176. doi: 10.1007/s00284-013-0338-1
|
| [4] |
Zhang X, Zhang D, Jia H, et al. (2015) The oral and gut microbiotas are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat Med 21: 895–905. doi: 10.1038/nm.3914
|
| [5] |
Chen J, Wright K, Davis JM, et al. (2016) An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med 8: 43–51. doi: 10.1186/s13073-016-0299-7
|
| [6] |
Sender R, Fuchs S, Milo R (2016) Revised estimates for the number of human and bacteria cells in the body. PLoS Biol 14: e1002533. doi: 10.1371/journal.pbio.1002533
|
| [7] |
Falony G, Joossens M, Vieira-Silva S, et al. (2016) Population-level analysis of gut microbiota variation. Science 352: 560–564. doi: 10.1126/science.aad3503
|
| [8] | Koppel N, Rekdal VM, Balskus EP (2017) Chemical transformation of xenobiotics by the human gut microbiota. Science 356: 1246–1257. |
| [9] |
Klatt NR, Cheu R, Birse K, et al. (2017) Vaginal bacteria modify HIV tenofovir microbicide efficacy in African women. Science 356: 938–945. doi: 10.1126/science.aai9383
|
| [10] |
Koh A, De Vadder F, Kovatcheva-Datchary P, et al. (2016) From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 165: 1332–1345. doi: 10.1016/j.cell.2016.05.041
|
| [11] | Takasuna K, Hagiwara T, Hirohashi M, et al. (1996) Involvement of b-glucuronidase in intestinal microflora in the intestinal toxicity of the antitumor camptothecin derivative irinotecan hydrochloride (CPT-11 ) in rats. Cancer Res 56: 3752–3757. |
| [12] |
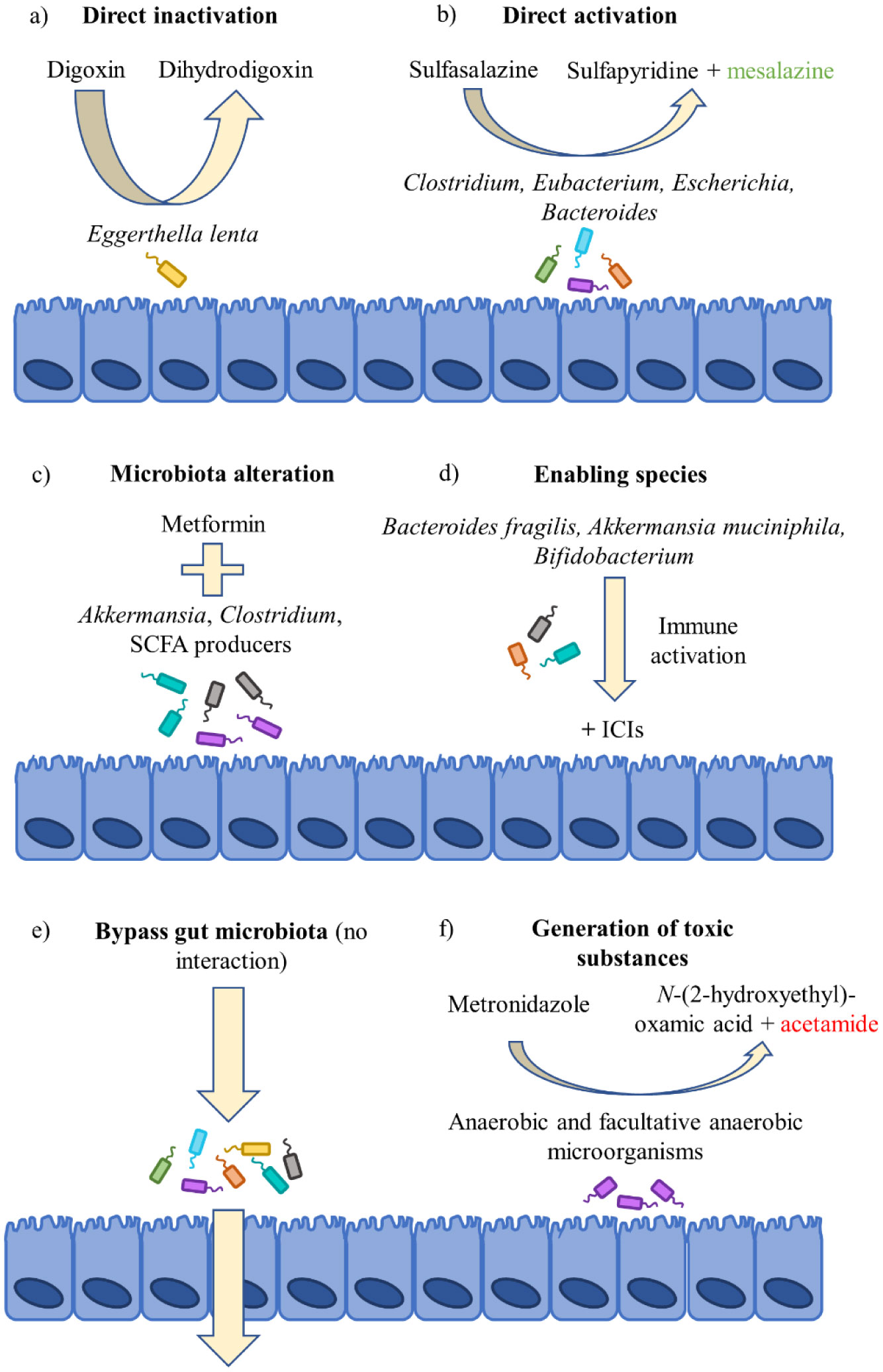
Lindenbaum J, Rund DG, Butler VP, et al. (1981) Inactivation of digoxin by the gut flora: Reversal by antibiotic therapy. N Engl J Med 305: 789–794. doi: 10.1056/NEJM198110013051403
|
| [13] | Dobkin J, Saha J, Butler V, et al. (1982) Digoxin-inactivating bacteria: Identification in human gut flora. Science 220: 325–327. |
| [14] |
Haiser HJ, Gootenberg DB, Chatman K, et al. (2013) Predicting and manipulating cardiac drug inactivation by the human gut bacterium Eggerthella lenta. Science 341: 295–298. doi: 10.1126/science.1235872
|
| [15] | Rafii F, Franklin W, Cerniglia CE (1990) Azoreductase activity of anaerobic bacteria isolated from human intestinal microflora. Appl Environ Microbiol 7: 2146–2151. |
| [16] | Rafii F, Cerniglia CE (1995) Reduction of azo dyes and nitroaromatic compounds by bacterial enzymes from the human intestinal tract. Environ Health Persp 103: 17–19. |
| [17] |
Azadkhan A, Truelove S, Aronson J (1982) The disposition and metabolism of sulphasalazine (salicylazosulphapyridine) in man. Brit J Clin Pharmaco 13: 523–528. doi: 10.1111/j.1365-2125.1982.tb01415.x
|
| [18] |
Bishop JB, Witt KL, Gulati DK, et al. (1990) Evaluation of the mutagenicity of the anti-inflammatory drug salicylazosulfapyridine (SASP). Mutagenesis 5: 549–554. doi: 10.1093/mutage/5.6.549
|
| [19] |
Das KM, Eastwood MA, McManus JPA, et al. (1973) Adverse reactions during salicylazosulfapyridine therapy and the relation with drug metabolism and acetylator phenotype. N Engl J Med 289: 491–495. doi: 10.1056/NEJM197309062891001
|
| [20] | Dingsdag SA, Hunter N (2017) Metronidazole: an update on metabolism, structure-cytotoxicity and resistance mechanisms. J Antimicrob Chemoth 73: 265–279. |
| [21] |
Bendesky A, Menéndez D, Ostrosky-Wegman P (2002) Is metronidazole carcinogenic? Mutat Res-Rev Mutat 511: 133–144. doi: 10.1016/S1383-5742(02)00007-8
|
| [22] |
Chrystal EJT, Koch RL, McLafferty MA, et al. (1980) Relationship between metronidazole metabolism and bactericidal activity. Antimicrob Agents Ch 18: 566–573. doi: 10.1128/AAC.18.4.566
|
| [23] |
Pardoll DM (2012) The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer 12: 252–264. doi: 10.1038/nrc3239
|
| [24] |
Dubin K, Callahan MK, Ren B, et al. (2016) Intestinal microbiota analyses identify melanoma patients at risk for checkpoint-blockade-induced colitis. Nat Commun 7: 10391–10397. doi: 10.1038/ncomms10391
|
| [25] |
Sivan A, Corrales L, Hubert N, et al. (2015) Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 350: 1084–1089. doi: 10.1126/science.aac4255
|
| [26] |
Routy B, Le Chatelier E, Derosa L, et al. (2018) Gut microbiota influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 359: 91–97. doi: 10.1126/science.aan3706
|
| [27] |
Matson V, Fessler J, Bao R, et al. (2018) The commensal microbiota is associated with anti- PD-1 efficacy in metastatic melanoma patients. Science 359: 104–108. doi: 10.1126/science.aao3290
|
| [28] |
Vétizou M, Pitt JM, Daillère R, et al. (2015) Anticancer immunotherapy by CTLA-4 blockade relies on the gutmicrobiota. Science 350: 1079–1084. doi: 10.1126/science.aad1329
|
| [29] | Shin NR, Lee JC, Lee HY, et al. (2014) An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 63: 727–735. |
| [30] |
Bonora E, Cigolini M, Bosello O, et al. (1984) Lack of effect of intravenous metformin on plasma concentrations of glucose, insulin, C-peptide, glucagon and growth hormone in non-diabetic subjects. Curr Med Res Opin 9: 47–51. doi: 10.1185/03007998409109558
|
| [31] |
Foretz M, Guigas B, Bertrand L, et al. (2014) Metformin: From mechanisms of action to therapies. Cell Metab 20: 953–966. doi: 10.1016/j.cmet.2014.09.018
|
| [32] |
Qin J, Li Y, Cai K, et al. (2012) A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 490: 55–60. doi: 10.1038/nature11450
|
| [33] |
Forslund K, Hildebrand F, Nielsen T, et al. (2015) Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature 528: 262–266. doi: 10.1038/nature15766
|
| [34] |
De La Cuesta-Zuluaga J, Mueller NT, Corrales-Agudelo V, et al. (2017) Metformin is associated with higher relative abundance of mucin-degrading akkermansia muciniphila and several short-chain fatty acid-producing microbiota in the gut. Diabetes Care 40: 54–62. doi: 10.2337/dc16-1324
|
| [35] |
Lee H (2014) Effect of metformin on metabolic improvement and gut microbiota. Appl Environ Microbiol 80: 5935–5943. doi: 10.1128/AEM.01357-14
|
| [36] |
Everard A, Belzer C, Geurts L, et al. (2013) Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. P Natl Acad Sci USA 110: 9066–9071. doi: 10.1073/pnas.1219451110
|
| [37] | Lie E, Uhlig T, van der Heijde D, et al. (201) Effectiveness of sulfasalazine and methotrexate in 1102 DMARD-nave patients with early RA. Rheumatology 51: 670–678. |
| [38] |
Wijbrandts CA, Tak PP (2017) Prediction of response to targeted treatment in rheumatoid arthritis. Mayo Clin Proc 92: 1129–1143. doi: 10.1016/j.mayocp.2017.05.009
|
| [39] |
Cutulo M, Sulli A, Pizzorni C, et al. (2001) Anti-inflammatory mechanisms of methotrexate in rheumatoid arthritis. Ann Rheum Dis 60: 729–735. doi: 10.1136/ard.60.8.729
|
| [40] |
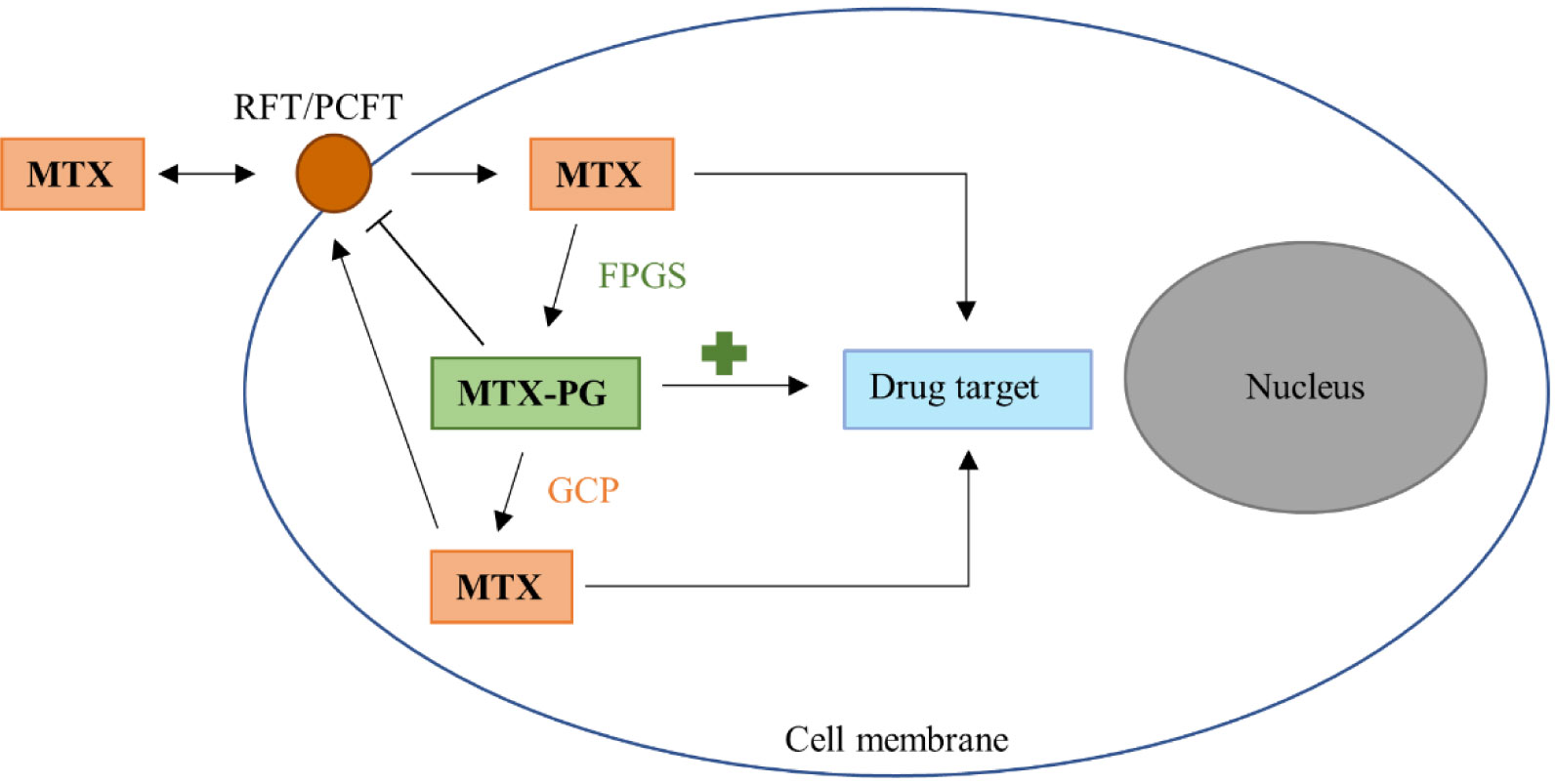
Takahashi C, Kaneko Y, Okano Y, et al. (2017) Association of erythrocyte methotrexate- polyglutamate levels with the efficacy and hepatotoxicity of methotrexate in patients with rheumatoid arthritis : a 76-week prospective study. RMD Open 3: e000363. doi: 10.1136/rmdopen-2016-000363
|
| [41] | Tian H, Cronstein BN (2007) Understanding the mechanisms of action of methotrexate-Implications for the treatment of rheumatoid arthritis. Bull NYU Hosp Jt Dis 65: 168–173. |
| [42] | Levy CC, Goldman P (1967) The enzymatic hydrolysis of methotrexate. J Biol Chem 242: 2933–2938. |
| [43] |
Webb M (1955) Inactivation of analogues of folic acid by certain non-exacting bacteria. Biochem Biophys Acta 17: 212–225. doi: 10.1016/0006-3002(55)90352-8
|
| [44] |
Larimer CM, Slavnic D, Pitstick LD, et al. (2014) Comparison of substrate specificity of Escherichia coli p-Aminobenzoyl-glutamate hydrolase with Pseudomonas carboxypeptidase G. Adv Enzym Res 2: 39–48. doi: 10.4236/aer.2014.21004
|
| [45] | Nayak R, O'Loughlin C, Fischbach M, et al. (2016) Methotrexate is an antibacterial drug metabolized by human gut bacteria-ACR Meeting Abstracts. Arthritis Rheumatol 68: 10. |
| [46] |
Dervieux T, Zablocki R, Kremer J (2010) Red blood cell methotrexate polyglutamates emerge as a function of dosage intensity and route of administration during pulse methotrexate therapy in rheumatoid arthritis. Rheumatology 49: 2337–2345. doi: 10.1093/rheumatology/keq216
|
| [47] |
Busquets D, Mas-de-Xaxars T, López-Siles M, et al. (2015) Anti-tumour necrosis factor treatment with adalimumab induces changes in the microbiota of Crohn's Disease. J Crohns Colitis 9: 899–906. doi: 10.1093/ecco-jcc/jjv119
|
| [48] |
Bazin T, Hooks KB, Barnetche T, et al. (2018) Microbiota composition may predict anti-TNF alpha response in spondyloarthritis patients: An exploratory study. Sci Rep 8: 5446–5449. doi: 10.1038/s41598-018-23571-4
|
| [49] |
Andersen V, Holmskov U, Sørensen SB, et al. (2017) Proposal for a study on treatment selection and lifestyle recommendations in chronic inflammatory diseases: A Danish multidisciplinary collaboration on prognostic factors and personalised medicine. Nutrients 9: 499–508. doi: 10.3390/nu9050499
|
Figures(2)

Ellie Sayers, Alex MacGregor, Simon R. Carding. Drug-microbiota interactions and treatment response: Relevance to rheumatoid arthritis[J]. AIMS Microbiology, 2018, 4(4): 642-654. doi: 10.3934/microbiol.2018.4.642







 DownLoad:
DownLoad: