Citation: Chayu Yang, Jin Wang. A cholera transmission model incorporating the impact of medical resources[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5226-5246. doi: 10.3934/mbe.2019261

| [1] | D. M. Hartley, J. G. Morris and D. L. Smith, Hyperinfectivity: A critical element in the ability of V. cholerae to cause epidemics? PLoS Med., 3 (2006), 0063–0069. |
| [2] | E. J. Nelson, J. B. Harris, J. G. Morris, et al., Cholera transmission: The host, pathogen and bacteriophage dynamics, Nat. Rev. Microbiol., 7 (2009), 693–702. |
| [3] | D. He, X. Wang, D. Gao, et al., Modeling the 2016-2017 Yemen cholera outbreak with the impact of limited medical resources, J. Theor. Biol., 451 (2018), 80–85. |
| [4] | A. Camacho, M. Bouhenia, R. Alyusfi, et al., Cholera epidemic in Yemen, 2016-18: an analysis of surveillance data, Lancet Glob. Health, published on line May 3, 2018. Available from: http: //doi.org/10.1016/S2214-109X(18)30230-4. |
| [5] | Z. Mukandavire, S. Liao, J. Wang, et al., Estimating the reproductive numbers for the 2008-2009 cholera outbreaks in Zimbabwe, P. Natl. Acad. Sci. USA, 108 (2011), 8767–8772. |
| [6] | A. R. Tuite, J. H. Tien, M. C. Eisenberg, et al., Cholera epidemic in Haiti, 2010 – using a trans-mission model to explain spatial spread of disease and identify optimal control interventions, Ann. Intern. Med., 154 (2011), 593–601. |
| [7] | P. R. Mason, Zimbabwe experiences the worst epidemic of cholera in Africa, J. Infect. Dev. Countr., 3 (2009), 148–151. |
| [8] | WHO cholera fact sheet, 20031 February 2018. Available from: http://www.who.int/en/news-room/fact-sheets/detail/cholera. |
| [9] | WHO report on global surveillance of epidemic-prone infectious diseases, 2000: 39-54. Available from: http://www.who.int/csr/resources/publications/surveillance/WHO_Report_Infectious_Diseases.pdf. |
| [10] | G. Zorlu, Cholera vaccine deployed to control African outbreak, Nature, News and Comment, June 2012. |
| [11] | S. Shin, S. N. Desai, B. K. Sah, et al., Oral vaccines against cholera, Clin. Infect. Dis., 52 (2011), 1343–1349. |
| [12] | S. Batterman, J. Eisenberg, R. Hardin, et al., Sustainable control of water-related infectious dis-eases: A review and proposal for interdisciplinary health-based systems research, Environ. Health Persp., 117 (2009), 1023–1032. |
| [13] | E. D. Mintz and R. V. Tauxe, Cholera in Africa: a closer look and a time for action, J. Infect. Dis., 208 (2013), S4–7. |
| [14] | S. Cumberland, An old enemy returns, B. World Health Organ., 87 (2009), 85–86. |
| [15] | D. Posny and J. Wang, Modeling cholera in periodic environments, J. Biol. Dynam., 8 (2014), 1–19. |
| [16] | J. H. Tien and D. J. Earn, Multiple transmission pathways and disease dynamics in a waterborne pathogen model, B. Math. Biol., 72 (2010), 1506–1533. |
| [17] | X. Wang, D. Posny and J. Wang, A reaction-convection-diffusion model for cholera spatial dy-namics, Discrete Cont. Dyn. B, 21 (2016), 2785–2809. |
| [18] | C. Yang, X. Wang, D. Gao, et al., Impact of awareness programs on cholera dynamics: Two modeling approaches, B. Math. Biol., 79 (2017), 2109–2131. |
| [19] | WHO survey on Yemen's health system. Available from: http://www.emro.who.int/media/news/survey-reveals-extent-of-damage-to-yemens-health-system.html. |
| [20] | WHO Weekly Epidemiology Bulletin, 21-27 May 2018. |
| [21] | WHO Yemen cholera situation reports. Available from: http://www.emro.who.int/yem/yemeninfocus/situation-reports.html. |
| [22] | R. M. Anderson and R. M. May, Infectious Diseases of Humans: Dynamics and Control, Oxford University Press, Oxford, 1991. |
| [23] | P. van den Driessche and J. Watmough, Reproduction number and subthreshold endemic equilibria for compartment models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [24] | O. Diekmann, J. A. P. Heesterbeek and A. J. Metz, On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous population, J. Math. Biol., 28 (1990), 365–382. |
| [25] | J. P. LaSalle, The stability of dynamical systems, Regional Conference Series in Applied Mathe-matics, SIAM, Philadelphia, 1976. |
| [26] | H. R. Thieme, Persistence under relaxed point-dissipativity (with application to an endemic model), SIAM J. Math. Anal., 24 (1993), 407–435. |
| [27] | D. Gao and S. Ruan, An SIS patch model with variable transmission coefficients, Math. Biosci., 232 (2011), 110–115. |
| [28] | M. Y. Li and J. S. Muldowney, Dynamics of differential equations on invariant manifolds, J. Differ. Equations, 168 (2000), 295–320. |
| [29] | M. Y. Li, J. S. Muldowney and P. V. D. Driessche, Global stability of SEIRS models in epidemiol-ogy, Canadian Appl. Math. Quart., 7 (1999), 409–425. |
| [30] | Wikipedia page for Yemen. Available from: http://en.wikipedia.org/wiki/Yemen. |
| [31] | WHO Global health observatory data repository: Life expectancy, 2013. Available from: http: //apps.who.int/gho/data/view.main.680?lang=en. |
| [32] | ClimateandAverageWeatherinYemen.Availablefrom: http://weather-and-climate.com/average-monthly-Rainfall-Temperature-Sunshine-in-Yemen. |
| [33] | World Climate Guide–Yemen. Available from: http://www.climatestotravel.com/climate/yemen. |
| [34] | A. A. King, E. L. Ionides, M. Pascual, et al., Inapparent infections and cholera dynamics, Nature, 454 (2008), 877–880. |
| [35] | X. Wang, X. Q. Zhao and J. Wang, A cholera epidemic model in a spatiotemporally heterogeneous environment, J. Math. Anal. Appl., 468 (2018), 893–912. |
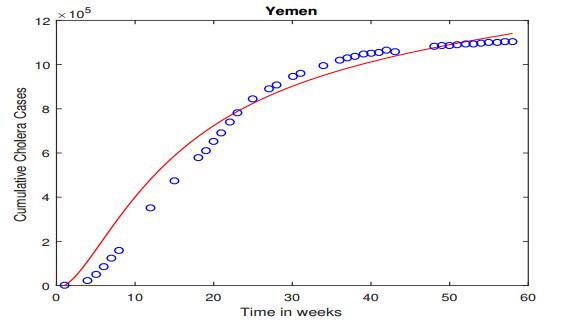
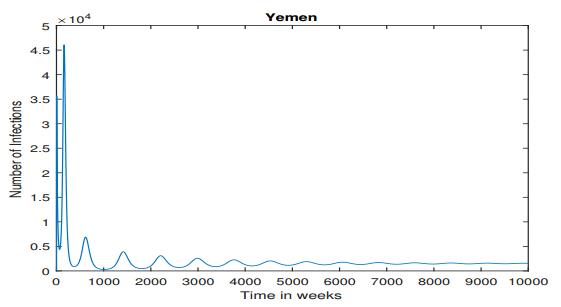
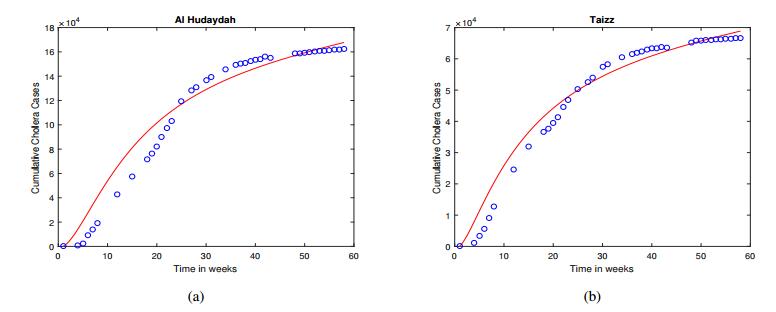
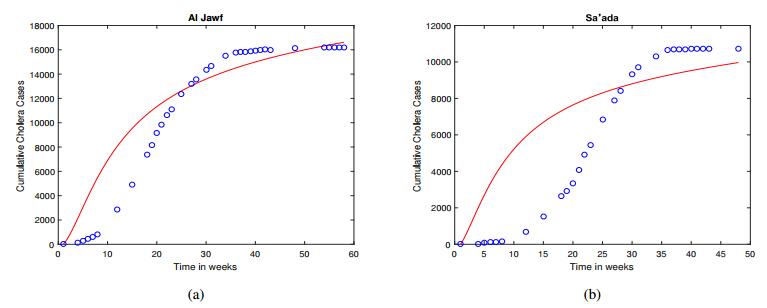
Figures(5) / Tables(6)

Chayu Yang, Jin Wang. A cholera transmission model incorporating the impact of medical resources[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5226-5246. doi: 10.3934/mbe.2019261







 DownLoad:
DownLoad: