Citation: Tin Phan, Changhan He, Alejandro Martinez, Yang Kuang. Dynamics and implications of models for intermittent androgen suppression therapy[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 187-204. doi: 10.3934/mbe.2019010

| [1] | Prostate cancer, Am. Cancer Soc., 4–18, 28–29, 56–57.0 |
| [2] | J. Baez and Y. Kuang, Mathematical models of androgen resistance in prostate cancer patients under intermittent androgen suppression therapy, Appl. Sci., 6 (2016), 352. |
| [3] | N. Bruchovsky, L. Klotz, J. Crook, S. Malone, C. Ludgate, W. J. Morris, M. E. Gleave and S. L. Goldenberg, Final results of the canadian prospective phase ii trial of intermittent androgen suppression for men in biochemical recurrence after radiotherapy for locally advanced prostate cancer, Cancer, 107 (2006), 389–395. |
| [4] | N. Buchan and S. Goldenberg, Intermittent versus continuous androgen suppression therapy: do we have consensus yet?, Cur. Oncol., 17 (2010), S45. |
| [5] | L. M. Cook, A. Araujo, J. M. Pow-Sang, M. M. Budzevich, D. Basanta and C. C. Lynch, Predictive computational modeling to define effective treatment strategies for bone metastatic prostate cancer, Sci. Rep., 6 (2016), 29384. |
| [6] | M. Droop, Some thoughts on nutrient limitation in algae, J. Phycol., 9 (1973), 264–272. |
| [7] | S. E. Eikenberry, J. D. Nagy and Y. Kuang, The evolutionary impact of androgen levels on prostate cancer in a multi-scale mathematical model, Biol. Direct, 5 (2010), 24. |
| [8] | M. C. Eisenberg and H. V. Jain, A confidence building exercise in data and identifiability: Modeling cancer chemotherapy as a case study, J. Theor. Biol., 431 (2017), 63–78. |
| [9] | M. Elishmereni, Y. Kheifetz, I. Shukrun, G. H. Bevan, D. Nandy, K. M. McKenzie, M. Kohli and Z. Agur, Predicting time to castration resistance in hormone sensitive prostate cancer by a personalization algorithm based on a mechanistic model integrating patient data, The Prostate, 76 (2016), 48–57. |
| [10] | R. Everett, A. Packer and Y. Kuang, Can mathematical models predict the outcomes of prostate cancer patients undergoing intermittent androgen deprivation therapy?, Biophys. Rev. Letters, 9 (2014), 173–191. |
| [11] | B. J. Feldman and D. Feldman, The development of androgen-independent prostate cancer, Nat. Rev. Cancer, 1 (2001), 34–45. |
| [12] | J. L. Fleck and C. G. Cassandras, Optimal design of personalized prostate cancer therapy using infinitesimal perturbation analysis, Nonlin. Analysis Hybrid System., 25 (2017), 246–262. |
| [13] | Q. Guo, Y. Tao and K. Aihara, Mathematical modeling of prostate tumor growth under intermittent androgen suppression with partial differential equations, lnt. J. Bifurcat. Chaos, 18 (2008), 3789– 3797. |
| [14] | T. Hatano, Y. Hirata, H. Suzuki and K. Aihara, Comparison between mathematical models of intermittent androgen suppression for prostate cancer, J. Theor. Biol., 366 (2015), 33–45. |
| [15] | Y. Hirata, N. Bruchovsky and K. Aihara, Development of a mathematical model that predicts the outcome of hormone therapy for prostate cancer, J. Theor. Biol., 264 (2010), 517–527. |
| [16] | Y. Hirata, K. Morino, K. Akakura, C. S. Higano and K. Aihara, Personalizing androgen suppression for prostate cancer using mathematical modeling, Sci. Rep., 8 (2018), 2673. |
| [17] | A. M. Ideta, G. Tanaka, T. Takeuchi and K. Aihara, A mathematical model of intermittent androgen suppression for prostate cancer, J. Nonlin. Sci., 18 (2008), 593–614. |
| [18] | T. L. Jackson, A mathematical model of prostate tumor growth and androgen-independent relapse, Discrete Cont. Dyn. B, 4 (2004), 187–202. |
| [19] | Y. Kuang, Basic properties of mathematical population models, J. Biomath., 17 (2002), 129–142. |
| [20] | Y. Kuang, J. D. Nagy and S. E. Eikenberry, Introduction to mathematical oncology, vol. 59, CRC Press, 2016. |
| [21] | J. D. Morken, A. Packer, R. A. Everett, J. D. Nagy and Y. Kuang, Mechanisms of resistance to intermittent androgen deprivation in patients with prostate cancer identified by a novel computational method, Cancer Res., 74 (2014), 3673–3683. |
| [22] | T. Portz, Y. Kuang and J. D. Nagy, A clinical data validated mathematical model of prostate cancer growth under intermittent androgen suppression therapy, Aip. Advances, 2 (2012), 011002. |
| [23] | A. Saltelli, K. Chan and E. M. Scott, Sensitivity analysis, Wiley New York, 2000. |
| [24] | T. Shimada and K. Aihara, A nonlinear model with competition between prostate tumor cells and its application to intermittent androgen suppression therapy of prostate cancer, Math. Biosci., 214 (2008), 134–139. |
| [25] | R. L. Siegel, K. D. Miller and A. Jemal, Cancer statistics, 2016, CA Cancer J. Clinic., 66 (2016), 7–30. |
| [26] | H. Vardhan Jain and A. Friedman, Modeling prostate cancer response to continuous versus intermittent androgen ablation therapy, Discrete Cont. Dyn. B, 18. |
| [27] | A. Wolf, R. C. Wender, R. B. Etzioni, I. M. Thompson, A. V. D'Amico, R. J. Volk, D. D. Brooks, C. Dash, I. Guessous, K. Andrews et al., American cancer society guideline for the early detection of prostate cancer: update 2010, CA Cancer J. Clinic., 60 (2010), 70–98. |
| [28] | J. Yang, T.J. Zhao, C.Q. Yuan, J.H. Xie and F.F. Hao, A nonlinear competitive model of the prostate tumor growth under intermittent androgen suppression, J. Theor. Biol., 404 (2016), 66– 72. |
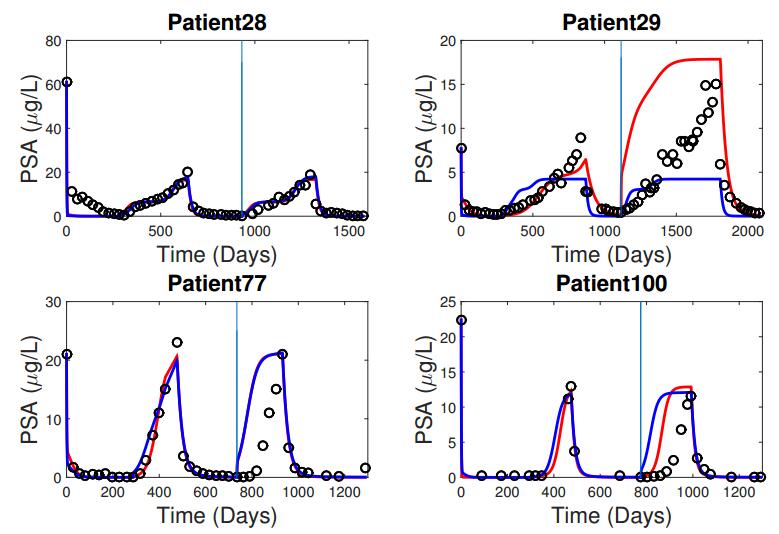
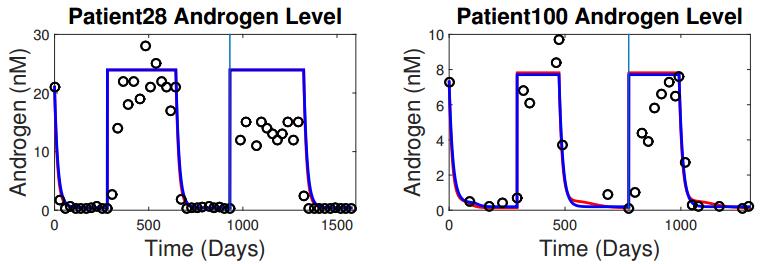
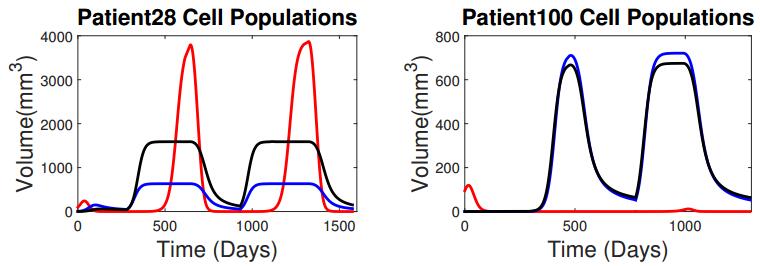
Figures(5) / Tables(3)

Tin Phan, Changhan He, Alejandro Martinez, Yang Kuang. Dynamics and implications of models for intermittent androgen suppression therapy[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 187-204. doi: 10.3934/mbe.2019010







 DownLoad:
DownLoad: