Citation: Yong Wang, Li-qun Gu. Biomedical diagnosis perspective of epigenetic detections using alpha-hemolysin nanopore[J]. AIMS Materials Science, 2015, 2(4): 448-472. doi: 10.3934/matersci.2015.4.448

| [1] |
Dupont C, Armant DR, Brenner CA (2009) Epigenetics: Definition, mechanisms and clinical perspective. Semin Reprod Med 27: 351-357. doi: 10.1055/s-0029-1237423

|
| [2] |
Dawson MA, Kouzarides T (2012) Cancer epigenetics: From mechanism to therapy. Cell 150: 12-27. doi: 10.1016/j.cell.2012.06.013
|
| [3] |
Esteller M (2008) Molecular origins of cancer: Epigenetics in cancer. New Engl J Med 358: 1148-1159+1096. doi: 10.1056/NEJMra072067
|
| [4] |
Laird PW (2005) Cancer epigenetics. Hum Mol Genet 14: R65-R76. doi: 10.1093/hmg/ddi113
|
| [5] |
Okugawa Y, Grady WM, Goel A (2015) Epigenetic Alterations in Colorectal Cancer: Emerging Biomarkers. Gastroenterology 149: 1204-1225e1212. doi: 10.1053/j.gastro.2015.07.011
|
| [6] | Deng D, Liu Z, Du Y (2010) Epigenetic Alterations as Cancer Diagnostic, Prognostic, and Predictive Biomarkers. Adv Genet 71: 126-176. |
| [7] |
Herceg Z, Hainaut P (2007) Genetic and epigenetic alterations as biomarkers for cancer detection, diagnosis and prognosis. Mol Oncol 1: 26-41. doi: 10.1016/j.molonc.2007.01.004
|
| [8] |
Dekker C (2007) Solid-state nanopores. Nat Nanotechnol 2: 209-215. doi: 10.1038/nnano.2007.27
|
| [9] |
Keyser UF (2011) Controlling molecular transport through nanopores. J R Soc Interface 8: 1369-1378. doi: 10.1098/rsif.2011.0222
|
| [10] |
Siwy ZS, Howorka S (2010) Engineered voltage-responsive nanopores. Chem Soc Rev 39: 1115-1132. doi: 10.1039/B909105J
|
| [11] |
Garaj S, Hubbard W, Reina A, et al. (2010) Graphene as a subnanometre trans-electrode membrane. Nature 467: 190-193. doi: 10.1038/nature09379
|
| [12] |
Siwy ZS, Davenport M (2010) Nanopores: Graphene opens up to DNA. Nat Nanotechnol 5: 697-698. doi: 10.1038/nnano.2010.198
|
| [13] |
Tada K, Haruyama J, Yang HX, et al. (2011) Graphene magnet realized by hydrogenated graphene nanopore arrays. Appl Phys Lett 99: 183111. doi: 10.1063/1.3653286
|
| [14] |
Ding S, Gao C, Gu LQ (2009) Capturing single molecules of immunoglobulin and ricin with an aptamer-encoded glass nanopore. Anal Chem 81: 6649-6655. doi: 10.1021/ac9006705
|
| [15] |
Gu LQ, Shim JW (2010) Single molecule sensing by nanopores and nanopore devices. Analyst 135: 441-451. doi: 10.1039/B907735A
|
| [16] | Fertig N, Meyer C, Blick RH, et al. (2001) Microstructured glass chip for ion-channel electrophysiology. Phys Rev E Stat Nonlin Soft Matter Phys 64: 409011-409014. |
| [17] |
Li J, Stein D, McMullan C, et al. (2001) Ion-beam sculpting at nanometre length scales. Nature 412: 166-169. doi: 10.1038/35084037
|
| [18] |
Storm AJ, Chen JH, Ling XS, et al. (2003) Fabrication of solid-state nanopores with single-nanometre precision. Nat Mater 2: 537-540. doi: 10.1038/nmat941
|
| [19] |
Kasianowicz JJ, Brandin E, Branton D, et al. (1996) Characterization of individual polynucleotide molecules using a membrane channel. Proc Natl Acad Sci U S A 93: 13770-13773. doi: 10.1073/pnas.93.24.13770
|
| [20] |
Laszlo AH, Derrington IM, Brinkerhoff H, et al. (2013) Detection and mapping of 5-methylcytosine and 5-hydroxymethylcytosine with nanopore MspA. Proc Natl Acad Sci U S A 110: 18904-18909. doi: 10.1073/pnas.1310240110
|
| [21] | Laszlo AH, Derrington IM, Ross BC, et al. (2014) Decoding long nanopore sequencing reads of natural DNA. Nat Biotechnol 32: 829-833. |
| [22] |
Wang Y, Montana V, Grubišić V, et al. (2015) Nanopore sensing of botulinum toxin type B by discriminating an enzymatically cleaved peptide from a synaptic protein synaptobrevin 2 derivative. ACS Appl Mater Inter 7: 184-192. doi: 10.1021/am5056596
|
| [23] |
Liu J, Eren E, Vijayaraghavan J, et al. (2012) OccK channels from Pseudomonas aeruginosa exhibit diverse single-channel electrical signatures but conserved anion selectivity. Biochemistry 51: 2319-2330. doi: 10.1021/bi300066w
|
| [24] |
Mohammad MM, Iyer R, Howard KR, et al. (2012) Engineering a rigid protein tunnel for biomolecular detection. J Am Chem Soc 134: 9521-9531. doi: 10.1021/ja3043646
|
| [25] |
Cheneke BR, van den Berg B, Movileanu L (2015) Quasithermodynamic contributions to the fluctuations of a protein nanopore. ACS Chem Biol 10: 784-794. doi: 10.1021/cb5008025
|
| [26] |
Wang HY, Li Y, Qin LX, et al. (2013) Single-molecule DNA detection using a novel SP1 protein nanopore. Chem Commun (Camb) 49: 1741-1743. doi: 10.1039/c3cc38939a
|
| [27] |
Wang S, Haque F, Rychahou PG, et al. (2013) Engineered nanopore of phi29 dna-packaging motor for real-time detection of single colon cancer specific antibody in serum. ACS Nano 7: 9814-9822. doi: 10.1021/nn404435v
|
| [28] |
Wu D, Bi S, Zhang L, et al. (2014) Single-molecule study of proteins by biological nanopore sensors. Sensors (Basel) 14: 18211-18222. doi: 10.3390/s141018211
|
| [29] |
Venkatesan BM, Bashir R (2011) Nanopore sensors for nucleic acid analysis. Nat Nanotechnol 6: 615-624. doi: 10.1038/nnano.2011.129
|
| [30] |
Hall AR, Scott A, Rotem D, et al. (2010) Hybrid pore formation by directed insertion of +Ý-haemolysin into solid-state nanopores. Nat Nanotechnol 5: 874-877. doi: 10.1038/nnano.2010.237
|
| [31] |
Bell NA, Engst CR, Ablay M, et al. (2012) DNA origami nanopores. Nano Lett 12: 512-517. doi: 10.1021/nl204098n
|
| [32] |
Iqbal SM, Akin D, Bashir R (2007) Solid-state nanopore channels with DNA selectivity. Nat Nanotechnol 2: 243-248. doi: 10.1038/nnano.2007.78
|
| [33] |
Tinazli A, Tang J, Valiokas R, et al. (2005) High-affinity chelator thiols for switchable and oriented immobilization of histidine-tagged proteins: a generic platform for protein chip technologies. Chemistry 11: 5249-5259. doi: 10.1002/chem.200500154
|
| [34] |
Wei R, Pedone D, Zurner A, et al. (2010) Fabrication of metallized nanopores in silicon nitride membranes for single-molecule sensing. Small 6: 1406-1414. doi: 10.1002/smll.201000253
|
| [35] |
Yusko EC, Johnson JM, Majd S, et al. (2011) Controlling protein translocation through nanopores with bio-inspired fluid walls. Nat Nanotechnol 6: 253-260. doi: 10.1038/nnano.2011.12
|
| [36] | Wang D, Harrer S, Luan B, et al. (2014) Regulating the transport of DNA through Biofriendly Nanochannels in a thin solid membrane. Sci Rep 4: 3985. |
| [37] | Branton D, Deamer DW, Marziali A, et al. (2008) The potential and challenges of nanopore sequencing. Nat Nanotechnol 26: 1146-1153. |
| [38] |
Cherf GM, Lieberman KR, Rashid H, et al. (2012) Automated forward and reverse ratcheting of DNA in a nanopore at 5-A precision. Nat Biotechnol 30: 344-348. doi: 10.1038/nbt.2147
|
| [39] |
Kasianowicz JJ, Brandin E, Branton D, et al. (1996) Characterization of individual polynucleotide molecules using a membrane channel. Proc Natl Acad Sci U S A 93: 13770-13773. doi: 10.1073/pnas.93.24.13770
|
| [40] |
Manrao EA, Derrington IM, Laszlo AH, et al. (2012) Reading DNA at single-nucleotide resolution with a mutant MspA nanopore and phi29 DNA polymerase. Nat Biotechnol 30: 349-353. doi: 10.1038/nbt.2171
|
| [41] |
An N, Fleming AM, White HS, et al. (2012) Crown ether-electrolyte interactions permit nanopore detection of individual DNA abasic sites in single molecules. Proc Natl Acad Sci U S A 109: 11504-11509. doi: 10.1073/pnas.1201669109
|
| [42] |
Wallace EV, Stoddart D, Heron AJ, et al. (2010) Identification of epigenetic DNA modifications with a protein nanopore. Chem Commun(Camb) 46: 8195-8197. doi: 10.1039/c0cc02864a
|
| [43] |
Shim J, Kim Y, Humphreys GI, et al. (2015) Nanopore-based assay for detection of methylation in double-stranded DNA fragments. ACS Nano 9: 290-300. doi: 10.1021/nn5045596
|
| [44] |
Bayley H, Cremer PS (2001) Stochastic sensors inspired by biology. Nature 413: 226-230. doi: 10.1038/35093038
|
| [45] |
Gu LQ, Shim JW (2010) Single molecule sensing by nanopores and nanopore devices. Analyst 135: 441-451. doi: 10.1039/B907735A
|
| [46] |
Howorka S, Siwy Z (2009) Nanopore analytics: sensing of single molecules. Chem Soc Rev 38: 2360-2384. doi: 10.1039/b813796j
|
| [47] | Luan B, Peng H, Polonsky S, et al. (2010) Base-by-base ratcheting of single stranded DNA through a solid-state nanopore. Phys Rev Lett 104. |
| [48] |
Asandei A, Chinappi M, Lee JK, et al. (2015) Placement of oppositely charged aminoacids at a polypeptide termini determines the voltage-controlled braking of polymer transport through nanometer-scale pores. Sci Rep 5: 10419. doi: 10.1038/srep10419
|
| [49] | Mereuta L, Roy M, Asandei A, et al. (2014) Slowing down single-molecule trafficking through a protein nanopore reveals intermediates for peptide translocation. Sci Rep 4: 3885. |
| [50] |
Deamer D (2010) Nanopore analysis of nucleic acids bound to exonucleases and polymerases. Annu Rev Biophys 39: 79-90. doi: 10.1146/annurev.biophys.093008.131250
|
| [51] |
Olasagasti F, Lieberman KR, Benner S, et al. (2010) Replication of individual DNA molecules under electronic control using a protein nanopore. Nat Nanotechnol 5: 798-806. doi: 10.1038/nnano.2010.177
|
| [52] |
Chu J, Gonzalez-Lopez M, Cockroft SL, et al. (2010) Real-time monitoring of DNA polymerase function and stepwise single-nucleotide DNA strand translocation through a protein nanopore. Angew Chem Int Ed Engl 49: 10106-10109. doi: 10.1002/anie.201005460
|
| [53] |
Benner S, Chen RJ, Wilson NA, et al. (2007) Sequence-specific detection of individual DNA polymerase complexes in real time using a nanopore. Nat Nanotechnol 2: 718-724. doi: 10.1038/nnano.2007.344
|
| [54] |
Asandei A, Iftemi S, Mereuta L, et al. (2014) Probing of various physiologically relevant metals: Amyloid-β peptide interactions with a lipid membrane-immobilized protein nanopore. J Membrane Biol 247: 523-530. doi: 10.1007/s00232-014-9662-z
|
| [55] |
Asandei A, Schiopu I, Iftemi S, et al. (2013) Investigation of Cu2+ binding to human and rat amyloid fragments Aβ (1-16) with a protein nanopore. Langmuir 29: 15634-15642. doi: 10.1021/la403915t
|
| [56] |
Ying YL, Wang HY, Sutherland TC, et al. (2011) Monitoring of an ATP-binding aptamer and its conformational changes using an α-hemolysin nanopore. Small 7: 87-94. doi: 10.1002/smll.201001428
|
| [57] |
Clarke J, Wu HC, Jayasinghe L, et al. (2009) Continuous base identification for single-molecule nanopore DNA sequencing. Nat Nanotechnol 4: 265-270. doi: 10.1038/nnano.2009.12
|
| [58] |
Branton D, Deamer DW, Marziali A, et al. (2008) The potential and challenges of nanopore sequencing. Nat Biotechnol 26: 1146-1153. doi: 10.1038/nbt.1495
|
| [59] |
Bayley H (2006) Sequencing single molecules of DNA. Curr Opin Chem Biol 10: 628-637. doi: 10.1016/j.cbpa.2006.10.040
|
| [60] |
Bartel DP (2004) MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 116: 281-297. doi: 10.1016/S0092-8674(04)00045-5
|
| [61] |
Carthew RW, Sontheimer EJ (2009) Origins and Mechanisms of miRNAs and siRNAs. Cell 136: 642-655. doi: 10.1016/j.cell.2009.01.035
|
| [62] |
Calin GA, Croce CM (2006) MicroRNA signatures in human cancers. Nat Rev Cancer 6: 857-866. doi: 10.1038/nrc1997
|
| [63] |
Chen X, Ba Y, Ma L, et al. (2008) Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res 18: 997-1006. doi: 10.1038/cr.2008.282
|
| [64] |
Mitchell PS, Parkin RK, Kroh EM, et al. (2008) Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A 105: 10513-10518. doi: 10.1073/pnas.0804549105
|
| [65] |
Wang Y, Zheng D, Tan Q, et al. (2011) Nanopore-based detection of circulating microRNAs in lung cancer patients. Nat Nanotechnol 6: 668-674. doi: 10.1038/nnano.2011.147
|
| [66] |
Tian K, He Z, Wang Y, et al. (2013) Designing a polycationic probe for simultaneous enrichment and detection of microRNAs in a nanopore. ACS Nano 7: 3962-3969. doi: 10.1021/nn305789z
|
| [67] |
Zhang X, Wang Y, Fricke BL, et al. (2014) Programming nanopore ion flow for encoded multiplex microRNA detection. ACS Nano 8: 3444-3450. doi: 10.1021/nn406339n
|
| [68] |
Cao C, Ying YL, Gu Z, et al. (2014) Enhanced resolution of low molecular weight poly(ethylene glycol) in nanopore analysis. Anal Chem 86: 11946-11950. doi: 10.1021/ac504233s
|
| [69] |
Baral A, Kumar P, Pathak R, et al. (2013) Emerging trends in G-quadruplex biology-role in epigenetic and evolutionary events. Mol BioSyst 9: 1568-1575. doi: 10.1039/c3mb25492e
|
| [70] |
Frees S, Menendez C, Crum M, et al. (2014) QGRS-Conserve: A computational method for discovering evolutionarily conserved G-quadruplex motifs. Human Genomics 8: 8. doi: 10.1186/1479-7364-8-8
|
| [71] |
Paeschke K, Bochman ML, Daniela Garcia P, et al. (2013) Pif1 family helicases suppress genome instability at G-quadruplex motifs. Nature 497: 458-462. doi: 10.1038/nature12149
|
| [72] |
Qiu J, Wang M, Zhang Y, et al. (2015) Biological function and medicinal research significance of g-quadruplex interactive proteins. Curr Top Med Chem 15: 1971-1987. doi: 10.2174/1568026615666150515150803
|
| [73] |
Fleming AM, Burrows CJ (2013) G-quadruplex folds of the human telomere sequence alter the site reactivity and reaction pathway of guanine oxidation compared to duplex DNA. Chem Res Toxicol 26: 593-607. doi: 10.1021/tx400028y
|
| [74] |
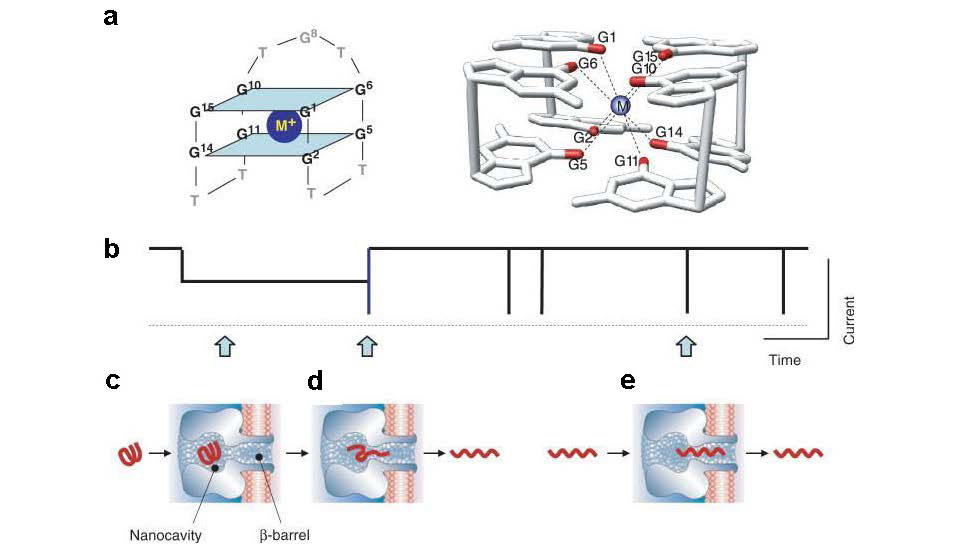
An N, Fleming AM, Middleton EG, et al. (2014) Single-molecule investigation of G-quadruplex folds of the human telomere sequence in a protein nanocavity. Proc Natl Acad Sci U S A 111: 14325-14331. doi: 10.1073/pnas.1415944111
|
| [75] |
An N, Fleming AM, Burrows CJ (2013) Interactions of the human telomere sequence with the nanocavity of the α-hemolysin ion channel reveal structure-dependent electrical signatures for hybrid folds. J Am Chem Soc 135: 8562-8570. doi: 10.1021/ja400973m
|
| [76] |
Wolna AH, Fleming AM, Burrows CJ (2014) Single-molecule analysis of thymine dimer-containing G-quadruplexes formed from the human telomere sequence. Biochemistry 53: 7484-7493. doi: 10.1021/bi501072m
|
| [77] |
Shim JW, Gu LQ (2008) Encapsulating a single G-quadruplex aptamer in a protein nanocavity. J Phys Chem B 112: 8354-8360. doi: 10.1021/jp0775911
|
| [78] |
Shim JW, Tan Q, Gu LQ (2009) Single-molecule detection of folding and unfolding of the G-quadruplex aptamer in a nanopore nanocavity. Nucleic Acids Res 37: 972-982. doi: 10.1093/nar/gkn968
|
| [79] |
Shim J, Gu LQ (2012) Single-molecule investigation of G-quadruplex using a nanopore sensor. Methods 57: 40-46. doi: 10.1016/j.ymeth.2012.03.026
|
| [80] | Yang J, Xu ZP, Huang Y, et al. (2004) ATM and ATR: Sensing DNA damage. World J Gastroentero 10: 155-160. |
| [81] |
Diculescu VC, Paquim AMC, Brett AMO (2005) Electrochemical DNA sensors for detection of DNA damage. Sensors 5: 377-393. doi: 10.3390/s5060377
|
| [82] |
Fojta M (2002) Electrochemical sensors for DNA interactions and damage. Electroanalysis 14: 1449-1463. doi: 10.1002/1521-4109(200211)14:21<1449::AID-ELAN1449>3.0.CO;2-Z
|
| [83] |
Paleček E, Bartošík M (2012) Electrochemistry of nucleic acids. Chem Rev 112: 3427-3481. doi: 10.1021/cr200303p
|
| [84] |
An N, Fleming AM, White HS, et al. (2015) Nanopore detection of 8-oxoguanine in the human telomere repeat sequence. ACS Nano 9: 4296-4307. doi: 10.1021/acsnano.5b00722
|
| [85] |
Boiteux S, Guillet M (2004) Abasic sites in DNA: Repair and biological consequences in Saccharomyces cerevisiae. DNA Repair 3: 1-12. doi: 10.1016/j.dnarep.2003.10.002
|
| [86] |
Wilson Iii DM, Barsky D (2001) The major human abasic endonuclease: Formation, consequences and repair of abasic lesions in DNA. Mutat Res-DNA Repair 485: 283-307. doi: 10.1016/S0921-8777(01)00063-5
|
| [87] | Reardon JT, Sancar A (2005) Nucleotide Excision Repair. In: Moldave K, editor. Progress in Nucleic Acid Research and Molecular Biology, 183-235. |
| [88] |
Gates KS (2009) An overview of chemical processes that damage cellular DNA: Spontaneous hydrolysis, alkylation, and reactions with radicals. Chem Res Toxicol 22: 1747-1760. doi: 10.1021/tx900242k
|
| [89] |
Greco NJ, Tor Y (2005) Simple fluorescent pyrimidine analogues detect the presence of DNA abasic sites. J Am Chem Soc 127: 10784-10785. doi: 10.1021/ja052000a
|
| [90] |
Lucotti A, Tommasini M, Pezzoli D, et al. (2014) Molecular interactions of DNA with transfectants: A study based on infrared spectroscopy and quantum chemistry as aids to fluorescence spectroscopy and dynamic light scattering analyses. RSC Adv 4: 49620-49627. doi: 10.1039/C4RA08845J
|
| [91] |
Sun HB, Qian L, Yokota H (2001) Detection of abasic sites on individual DNA molecules using atomic force microscopy. Anal Chem 73: 2229-2232. doi: 10.1021/ac0013249
|
| [92] |
Bowler FR, Diaz-Mochon JJ, Swift MD, et al. (2010) DNA analysis by dynamic chemistry. Angew Chem Int Edit 49: 1809-1812. doi: 10.1002/anie.200905699
|
| [93] |
Atamna H, Cheung I, Ames BN (2000) A method for detecting abasic sites in living cells: Age-dependent changes in base excision repair. Proc Natl Acad Sci U S A 97: 686-691. doi: 10.1073/pnas.97.2.686
|
| [94] |
Kojima N, Takebayashi T, Mikami A, et al. (2009) Construction of highly reactive probes for abasic site detection by introduction of an aromatic and a guanidine residue into an aminooxy group. J Am Chem Soc 131: 13208-13209. doi: 10.1021/ja904767k
|
| [95] |
Zeglis BM, Boland JA, Barton JK (2008) Targeting abasic sites and single base bulges in DNA with metalloinsertors. J Am Chem Soc 130: 7530-7531. doi: 10.1021/ja801479y
|
| [96] |
Wang Y, Liu L, Wu C, et al. (2009) Direct detection and quantification of abasic sites for in vivo studies of DNA damage and repair. Nucl Med Biol 36: 975-983. doi: 10.1016/j.nucmedbio.2009.07.007
|
| [97] |
Hada M, Sutherland BM (2006) Spectrum of complex DNA damages depends on the incident radiation. Radiat Res 165: 223-230. doi: 10.1667/RR3498.1
|
| [98] |
An N, Fleming AM, White HS, et al. (2012) Crown ether-electrolyte interactions permit nanopore detection of individual DNA abasic sites in single molecules. Proc Natl Acad Sci U S A 109: 11504-11509. doi: 10.1073/pnas.1201669109
|
| [99] |
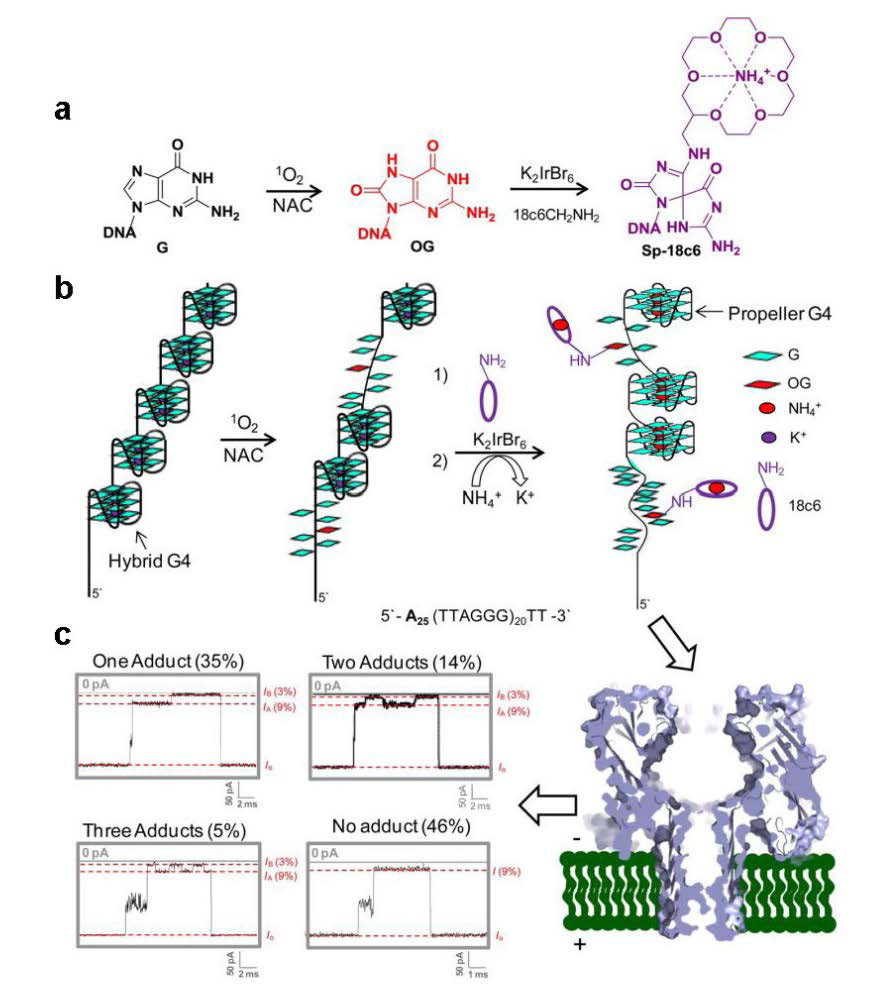
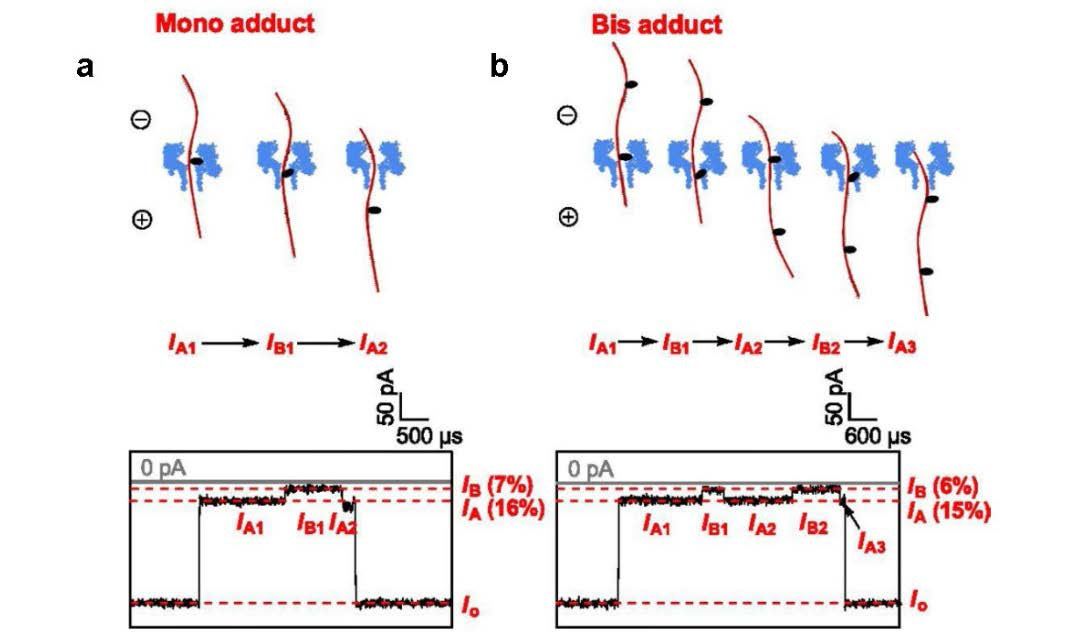
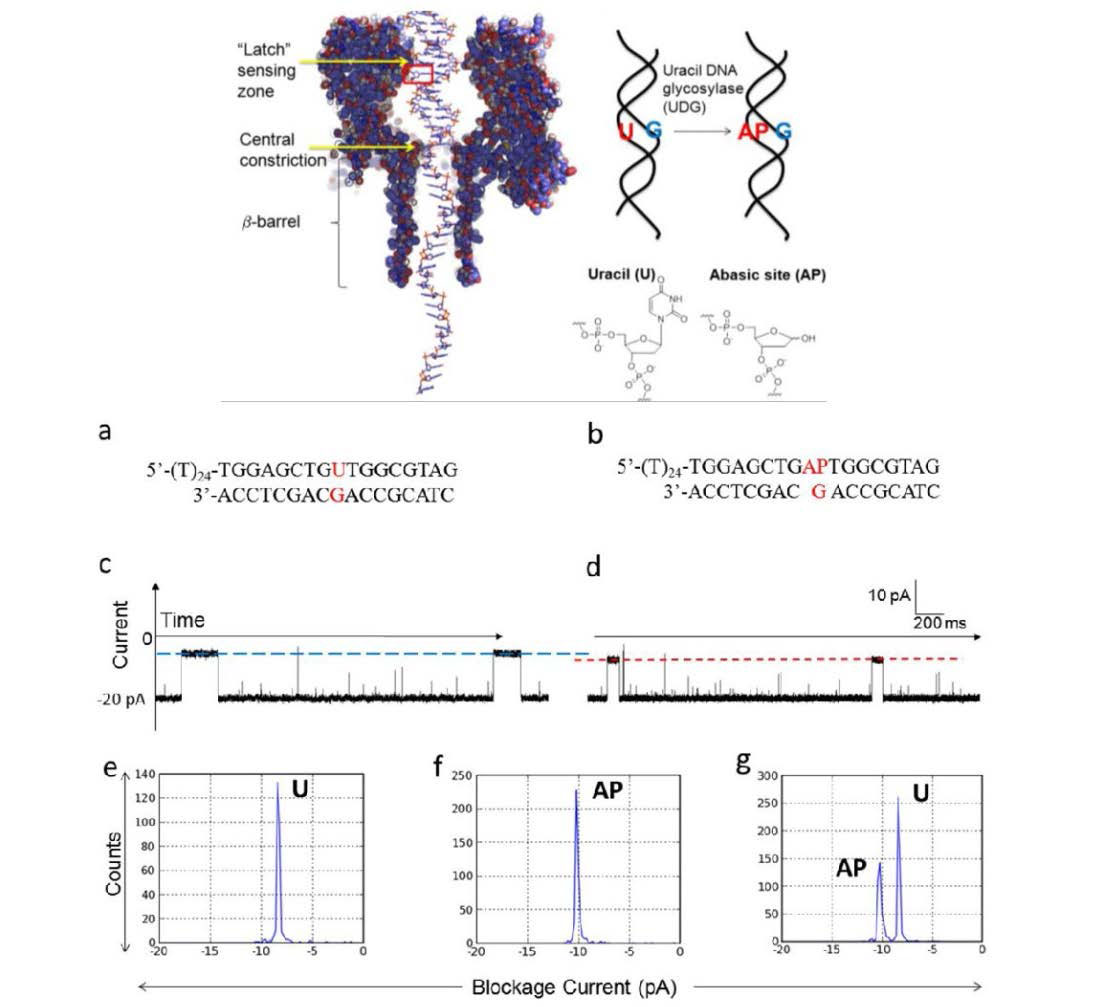
Jin Q, Fleming AM, Johnson RP, et al. (2013) Base-excision repair activity of uracil-DNA glycosylase monitored using the latch zone of α-hemolysin. J Am Chem Soc 135: 19347-19353. doi: 10.1021/ja410615d
|
| [100] |
Johnson RP, Fleming AM, Jin Q, et al. (2014) Temperature and electrolyte optimization of the α-hemolysin latch sensing zone for detection of base modification in double-stranded DNA. Biophys J 107: 924-931. doi: 10.1016/j.bpj.2014.07.006
|
| [101] | Lin YH, Tseng WL (2009) Highly sensitive and selective detection of silver ions and silver nanoparticles in aqueous solution using an oligonucleotide-based fluorogenic probe. Chem Commun (Camb): 6619-6621. |
| [102] | Ono A, Cao S, Togashi H, et al. (2008) Specific interactions between silver(I) ions and cytosine-cytosine pairs in DNA duplexes. Chem Commun (Camb): 4825-4827. |
| [103] |
Wen Y, Xing F, He S, et al. (2010) A graphene-based fluorescent nanoprobe for silver(I) ions detection by using graphene oxide and a silver-specific oligonucleotide. Chem Commun (Camb) 46: 2596-2598. doi: 10.1039/b924832c
|
| [104] |
Torigoe H, Okamoto I, Dairaku T, et al. (2012) Thermodynamic and structural properties of the specific binding between Ag(+) ion and C:C mismatched base pair in duplex DNA to form C-Ag-C metal-mediated base pair. Biochimie 94: 2431-2440. doi: 10.1016/j.biochi.2012.06.024
|
| [105] |
Miyake Y, Togashi H, Tashiro M, et al. (2006) MercuryII-mediated formation of thymine-HgII-thymine base pairs in DNA duplexes. J Am Chem Soc 128: 2172-2173. doi: 10.1021/ja056354d
|
| [106] |
Tanaka Y, Oda S, Yamaguchi H, et al. (2007) 15N-15N J-coupling across Hg(II): direct observation of Hg(II)-mediated T-T base pairs in a DNA duplex. J Am Chem Soc 129: 244-245. doi: 10.1021/ja065552h
|
| [107] |
Torigoe H, Ono A, Kozasa T (2010) Hg(II) ion specifically binds with T:T mismatched base pair in duplex DNA. Chemistry 16: 13218-13225. doi: 10.1002/chem.201001171
|
| [108] |
Wen S, Zeng T, Liu L, et al. (2011) Highly sensitive and selective DNA-based detection of mercury(II) with alpha-hemolysin nanopore. J Am Chem Soc 133: 18312-18317. doi: 10.1021/ja206983z
|
| [109] |
Ono A, Torigoe H, Tanaka Y, et al. (2011) Binding of metal ions by pyrimidine base pairs in DNA duplexes. Chem Soc Rev 40: 5855-5866. doi: 10.1039/c1cs15149e
|
| [110] |
Brena RM, Huang TH, Plass C (2006) Quantitative assessment of DNA methylation: Potential applications for disease diagnosis, classification, and prognosis in clinical settings. J Mol Med (Berl) 84: 365-377. doi: 10.1007/s00109-005-0034-0
|
| [111] |
Kriaucionis S, Heintz N (2009) The nuclear DNA base 5-hydroxymethylcytosine is present in Purkinje neurons and the brain. Science 324: 929-930. doi: 10.1126/science.1169786
|
| [112] |
Lister R, Pelizzola M, Dowen RH, et al. (2009) Human DNA methylomes at base resolution show widespread epigenomic differences. Nature 462: 315-322. doi: 10.1038/nature08514
|
| [113] | Li WW, Gong L, Bayley H (2013) Single-molecule detection of 5-hydroxymethylcytosine in DNA through chemical modification and nanopore analysis. Angew Chem Int Ed 52: 4350-4355. |
| [114] | Shim J, Humphreys GI, Venkatesan BM, et al. (2013) Detection and quantification of methylation in DNA using solid-state nanopores. Sci Rep 3: 1389. |
| [115] |
Wallace EV, Stoddart D, Heron AJ, et al. (2010) Identification of epigenetic DNA modifications with a protein nanopore. Chem Commun (Camb) 46: 8195-8197. doi: 10.1039/c0cc02864a
|
| [116] |
Wanunu M, Cohen-Karni D, Johnson RR, et al. (2011) Discrimination of methylcytosine from hydroxymethylcytosine in DNA molecules. J Am Chem Soc 133: 486-492. doi: 10.1021/ja107836t
|
| [117] |
Yu M, Hon GC, Szulwach KE, et al. (2012) Tet-assisted bisulfite sequencing of 5-hydroxymethylcytosine. Nat Protoc 7: 2159-2170. doi: 10.1038/nprot.2012.137
|
| [118] |
Yu M, Hon GC, Szulwach KE, et al. (2012) Base-resolution analysis of 5-hydroxymethylcytosine in the mammalian genome. Cell 149: 1368-1380. doi: 10.1016/j.cell.2012.04.027
|
| [119] |
Booth MJ, Branco MR, Ficz G, et al. (2012) Quantitative sequencing of 5-methylcytosine and 5-hydroxymethylcytosine at single-base resolution. Science 336: 934-937. doi: 10.1126/science.1220671
|
| [120] | Wang Y, Luan BQ, Yang Z, et al. (2014) Single Molecule Investigation of Ag+ Interactions with Single Cytosine-, Methylcytosine- and Hydroxymethylcytosine-Cytosine Mismatches in a Nanopore. Sci Rep 4: 5883. |
| [121] |
Torigoe H, Miyakawa Y, Ono A, et al. (2011) Thermodynamic properties of the specific binding between Ag+ ions and C:C mismatched base pairs in duplex DNA. Nucleos Nucleot Nucl 30: 149-167. doi: 10.1080/15257770.2011.553210
|
| [122] |
Kistenmacher TJ, Rossi M, Marzilli LG (1979) Crystal and molecular structure of (nitrato)(1-methylcytosine)silver(I): An unusual cross-linked polymer containing a heavy metal and a modified nucleic acid constituent. Inorg Chem 18: 240-244. doi: 10.1021/ic50192a007
|
| [123] |
Urata H, Yamaguchi E, Nakamura Y, et al. (2011) Pyrimidine-pyrimidine base pairs stabilized by silver(I) ions. Chem Commun (Camb) 47: 941-943. doi: 10.1039/C0CC04091F
|
| [124] |
Wang Y, Ritzo B, Gu L-Q (2015) Silver (i) ions modulate the stability of DNA duplexes containing cytosine, methylcytosine and hydroxymethylcytosine at different salt concentrations. RSC Adv 5: 2655-2658. doi: 10.1039/C4RA14490B
|
| [125] |
Egger G, Liang G, Aparicio A, et al. (2004) Epigenetics in human disease and prospects for epigenetic therapy. Nature 429: 457-463. doi: 10.1038/nature02625
|
| [126] |
Bestor TH (2000) The DNA methyltransferases of mammals. Hum Mol Genet 9: 2395-2402. doi: 10.1093/hmg/9.16.2395
|
| [127] | Kang I, Wang Y, Reagan C, et al. (2013) Designing DNA interstrand lock for locus-specific methylation detection in a nanopore. Sci Rep 3: 2381. |
| [128] |
Calin GA, Croce CM (2006) MicroRNA signatures in human cancers. Nat Rev Cancer 6: 857-866. doi: 10.1038/nrc1997
|
| [129] |
Vlassov VV, Laktionov PP, Rykova EY (2010) Circulating nucleic acids as a potential source for cancer biomarkers. Curr Mol Med 10: 142-165. doi: 10.2174/156652410790963295
|
| [130] |
Mitchell PS, Parkin RK, Kroh EM, et al. (2008) Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl Acad Sci U S A 105: 10513-10518. doi: 10.1073/pnas.0804549105
|
| [131] | Oremek GM, Sapoutzis N (2003) Pro-gastrin-releasing peptide (Pro-GRP), a tumor marker for small cell lung cancer. Anticancer Res 23: 895-898. |
| [132] |
Molina R, Auge JM, Bosch X, et al. (2009) Usefulness of serum tumor markers, including progastrin-releasing peptide, in patients with lung cancer: correlation with histology. Tumour Biol 30: 121-129. doi: 10.1159/000224628
|
| [133] |
Albrethsen J, Moller CH, Olsen J, et al. (2006) Human neutrophil peptides 1, 2 and 3 are biochemical markers for metastatic colorectal cancer. Eur J Cancer 42: 3057-3064. doi: 10.1016/j.ejca.2006.05.039
|
| [134] |
Bassani-Sternberg M, Barnea E, Beer I, et al. (2010) Soluble plasma HLA peptidome as a potential source for cancer biomarkers. Proc Natl Acad Sci U S A 107: 18769-18776. doi: 10.1073/pnas.1008501107
|
| [135] |
Stella Tsai CS, Chen HC, Tung JN, et al. (2010) Serum cellular apoptosis susceptibility protein is a potential prognostic marker for metastatic colorectal cancer. Am J Pathol 176: 1619-1628. doi: 10.2353/ajpath.2010.090467
|
| [136] |
Stoeva SI, Lee JS, Smith JE, et al. (2006) Multiplexed detection of protein cancer markers with biobarcoded nanoparticle probes. J Am Chem Soc 128: 8378-8379. doi: 10.1021/ja0613106
|
| [137] |
Zheng W, He L (2010) Multiplexed detection of protein cancer markers on Au/Ag-barcoded nanorods using fluorescent-conjugated polymers. Anal Bioanal Chem 397: 2261-2270. doi: 10.1007/s00216-010-3834-1
|
| [138] | Jin H, Liu GL (2012) Fabrication and optical characterization of light trapping silicon nanopore and nanoscrew devices. Nanotechnology 23. |
| [139] | Merchant CA, Drndić M (2012) Graphene nanopore devices for DNA sensing. In: Gracheva ME, editor. Methods in Molecular Biology, 211-226. |
| [140] |
Park S, Lim J, Pak YE, et al. (2013) A solid state nanopore device for investigating the magnetic properties of magnetic nanoparticles. Sensors (Basel, Switzerland) 13: 6900-6909. doi: 10.3390/s130606900
|
| [141] |
Wu GS, Zhang Y, Si W, et al. (2014) Integrated solid-state nanopore devices for third generation DNA sequencing. Sci China Technol Sc 57: 1925-1935. doi: 10.1007/s11431-014-5644-8
|
| [142] | Inoue K, Kawano R, Yasuga H, et al. Logic operation in DNA nano device: Electrical input/output through biological nanopore; 2013; Freiburg. Chemical and Biological Microsystems Society. 1881-1883. |
| [143] |
Lim MC, Lee MH, Kim KB, et al. (2015) A mask-free passivation process for low noise nanopore devices. J Nanosci Nanotechno 15: 5971-5977. doi: 10.1166/jnn.2015.10500
|
| [144] |
Mulero R, Prabhu AS, Freedman KJ, et al. (2010) Nanopore-Based Devices for Bioanalytical Applications. J Assoc Lab Automat 15: 243-252. doi: 10.1016/j.jala.2010.01.009
|
| [145] |
Cockroft SL, Chu J, Amorin M, et al. (2008) A single-molecule nanopore device detects DNA polymerase activity with single-nucleotide resolution. J Am Chem Soc 130: 818-820. doi: 10.1021/ja077082c
|
| [146] |
Kawano R, Osaki T, Sasaki H, et al. (2010) A polymer-based nanopore-integrated microfluidic device for generating stable bilayer lipid membranes. Small 6: 2100-2104. doi: 10.1002/smll.201000997
|
| [147] |
Shim JW, Gu LQ (2007) Stochastic sensing on a modular chip containing a single-ion channel. Anal Chem 79: 2207-2213. doi: 10.1021/ac0614285
|
| [148] |
Kang XF, Cheley S, Rice-Ficht AC, et al. (2007) A storable encapsulated bilayer chip containing a single protein nanopore. J Am Chem Soc 129: 4701-4705. doi: 10.1021/ja068654g
|
| [149] |
Eisenstein M (2012) Oxford Nanopore announcement sets sequencing sector abuzz. Nat Biotechnol 30: 295-296. doi: 10.1038/nbt0412-295
|
| [150] |
Quick J, Quinlan AR, Loman NJ (2014) A reference bacterial genome dataset generated on the MinIONTM portable single-molecule nanopore sequencer. GigaScience 3: 22. doi: 10.1186/2047-217X-3-22
|
| [151] | Ashton PM, Nair S, Dallman T, et al. (2015) MinION nanopore sequencing identifies the position and structure of a bacterial antibiotic resistance island. Nat Biotechnol 33: 296-302. |
| [152] | Madoui MA, Engelen S, Cruaud C, et al. (2015) Genome assembly using Nanopore-guided long and error-free DNA reads. BMC Genomics 16. |
| [153] |
Loman NJ, Quick J, Simpson JT (2015) A complete bacterial genome assembled de novo using only nanopore sequencing data. Nat Methods 12: 733-735. doi: 10.1038/nmeth.3444
|
| [154] | Palshikar P, Sharma A, Chauhan CS, et al. (2013) Biophotonics: A novel approach biomedical diagnosis. Int J Pharm Sci Rev Res 21: 350-354. |
| [155] | Liu A, Ye B (2013) Application of gold nanoparticles in biomedical researches and diagnosis. Clin Lab 59: 23-36. |
| [156] |
Nahar J, Imam T, Tickle KS, et al. (2012) Computational intelligence for microarray data and biomedical image analysis for the early diagnosis of breast cancer. Expert Syst Appl 39: 12371-12377. doi: 10.1016/j.eswa.2012.04.045
|
| [157] |
Kang JW, Lue N, Kong CR, et al. (2011) Combined confocal Raman and quantitative phase microscopy system for biomedical diagnosis. Biomed Opt Express 2: 2484-2492. doi: 10.1364/BOE.2.002484
|
| [158] |
Chen KI, Li BR, Chen YT (2011) Silicon nanowire field-effect transistor-based biosensors for biomedical diagnosis and cellular recording investigation. Nano Today 6: 131-154. doi: 10.1016/j.nantod.2011.02.001
|
| [159] |
Balas C (2009) Review of biomedical optical imaging - A powerful, non-invasive, non-ionizing technology for improving in vivo diagnosis. Meas Sci Technol 20: 104020. doi: 10.1088/0957-0233/20/10/104020
|
| [160] |
Ellis DI, Goodacre R (2006) Metabolic fingerprinting in disease diagnosis: Biomedical applications of infrared and Raman spectroscopy. Analyst 131: 875-885. doi: 10.1039/b602376m
|
| [161] |
Bottiroli G, Croce AC (2004) Autofluorescence spectroscopy of cells and tissues as a tool for biomedical diagnosis. Photoch Photobio Sci 3: 189-210. doi: 10.1039/b310627f
|
| [162] | Urbanova M, Plzak J, Strnad H, et al. (2010) Circulating nucleic acids as a new diagnostic tool. Cell Mol Biol Lett 15: 242-259. |
| [163] |
Liu KJ, Brock MV, Shih Ie M, et al. (2010) Decoding circulating nucleic acids in human serum using microfluidic single molecule spectroscopy. J Am Chem Soc 132: 5793-5798. doi: 10.1021/ja100342q
|
| [164] |
Thompson L, Turko I, Murad F (2006) Mass spectrometry-based relative quantification of human neutrophil peptides 1, 2, and 3 from biological samples. Mol Immunol 43: 1485-1489. doi: 10.1016/j.molimm.2005.08.001
|
| [165] |
Wanunu M, Dadosh T, Ray V, et al. (2010) Rapid electronic detection of probe-specific microRNAs using thin nanopore sensors. Nat Nanotechnol 5: 807-814. doi: 10.1038/nnano.2010.202
|
| [166] |
Squires A, Meller A (2013) DNA Capture and Translocation through Nanoscale Pores-a Fine Balance of Electrophoresis and Electroosmosis. Biophys J 105: 543-544. doi: 10.1016/j.bpj.2013.06.008
|
| [167] |
Wanunu M, Morrison W, Rabin Y, et al. (2010) Electrostatic focusing of unlabelled DNA into nanoscale pores using a salt gradient. Nat Nanotechnol 5: 160-165. doi: 10.1038/nnano.2009.379
|
| [168] |
Hatlo MM, Panja D, van Roij R (2011) Translocation of DNA molecules through nanopores with salt gradients: the role of osmotic flow. Phys Rev Lett 107: 068101. doi: 10.1103/PhysRevLett.107.068101
|
| [169] |
He Y, Tsutsui M, Scheicher RH, et al. (2013) Mechanism of How Salt-Gradient-Induced Charges Affect the Translocation of DNA Molecules through a Nanopore. Biophys J 105: 776-782. doi: 10.1016/j.bpj.2013.05.065
|
| [170] |
Chou T (2009) Enhancement of charged macromolecule capture by nanopores in a salt gradient. J Chem Phys 131: 034703. doi: 10.1063/1.3170952
|
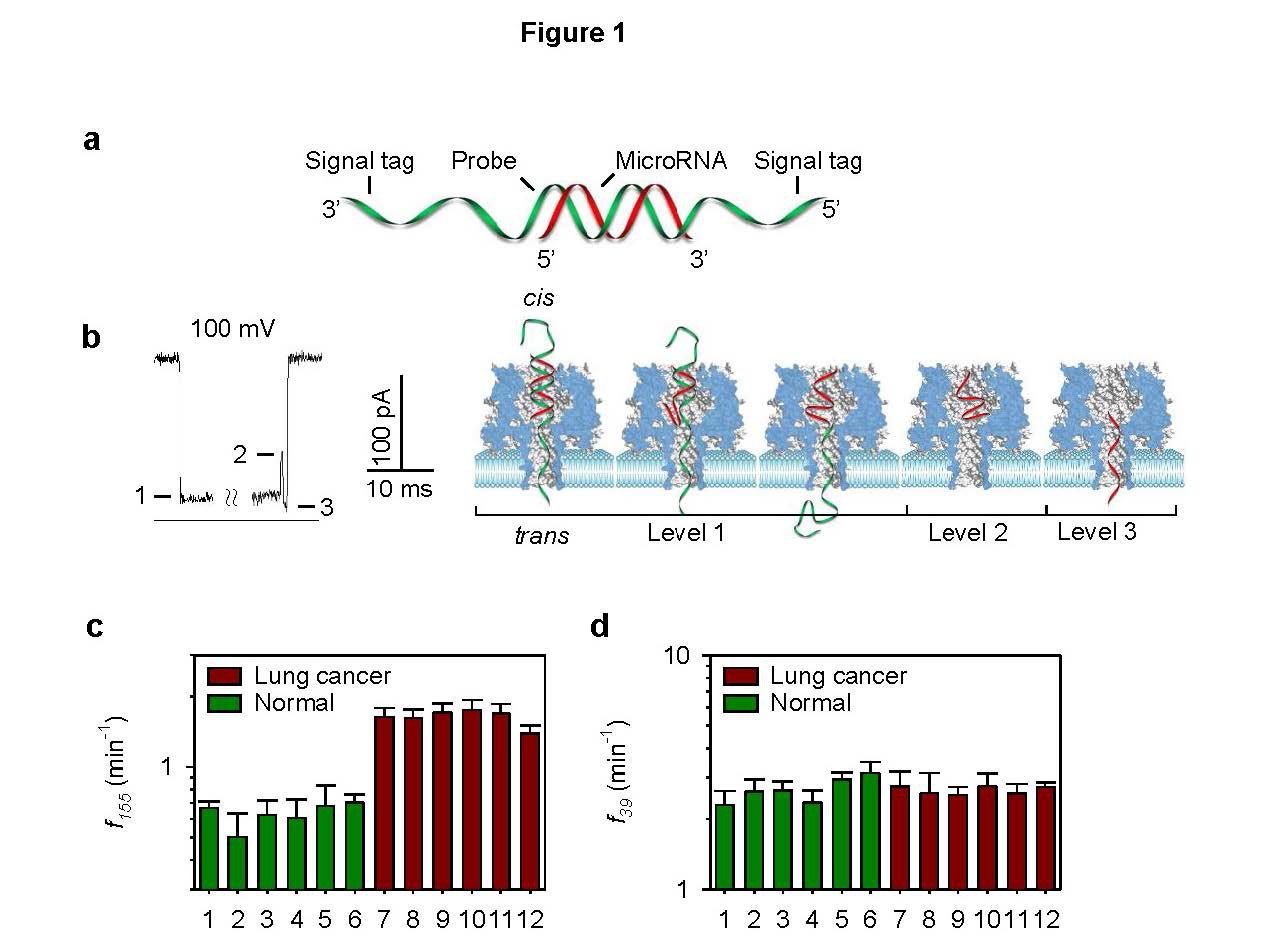
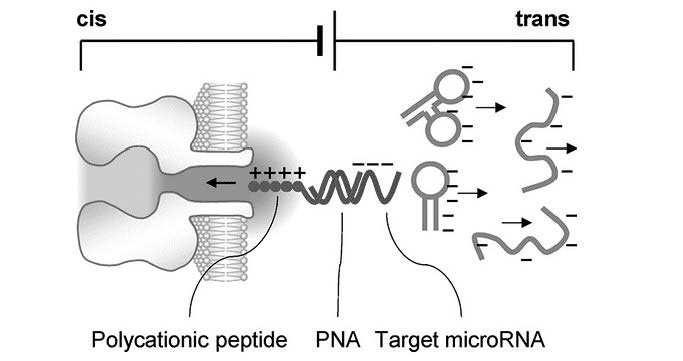
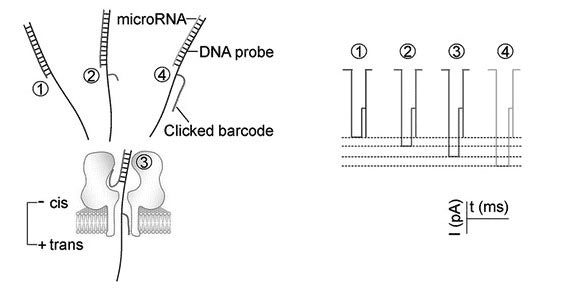
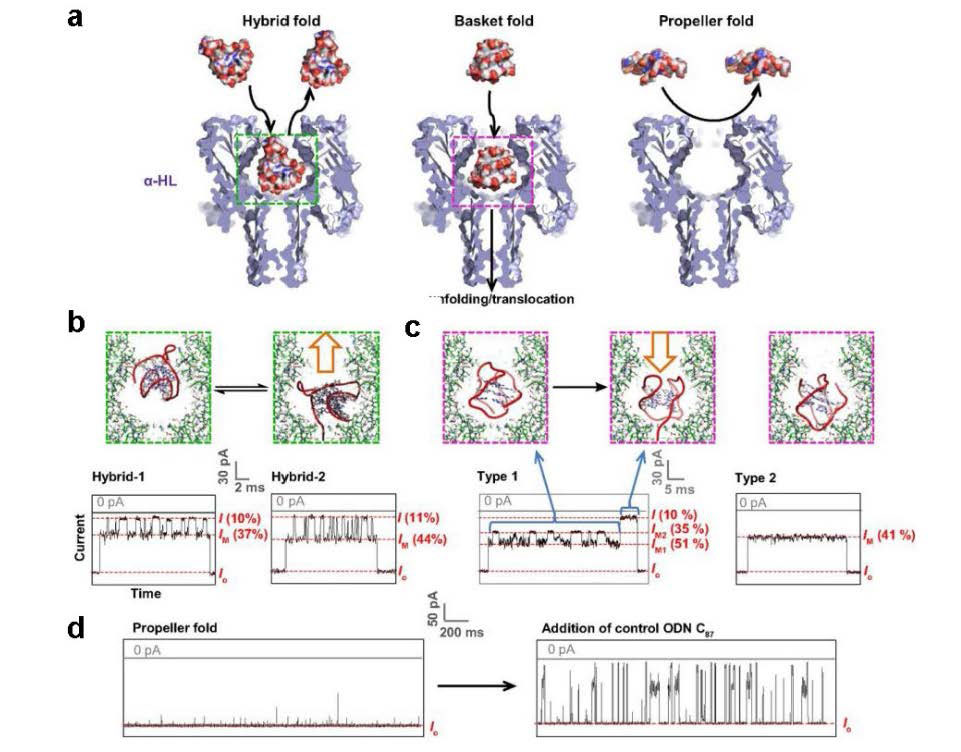
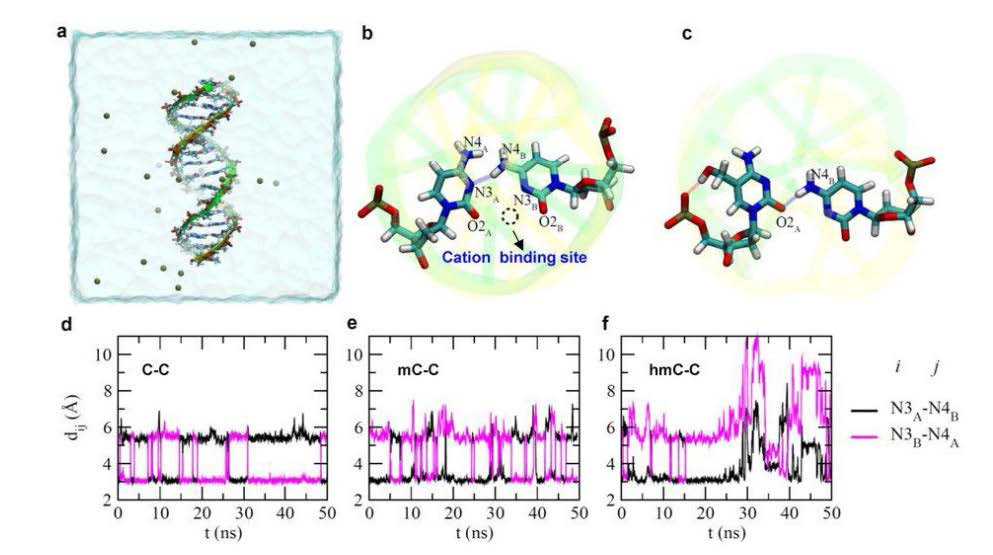
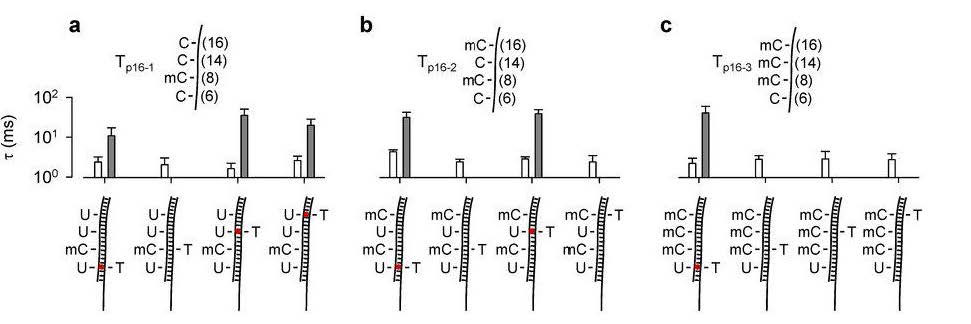
Figures(10)

Yong Wang, Li-qun Gu. Biomedical diagnosis perspective of epigenetic detections using alpha-hemolysin nanopore[J]. AIMS Materials Science, 2015, 2(4): 448-472. doi: 10.3934/matersci.2015.4.448







 DownLoad:
DownLoad: