Citation: Jana Söderlund, Peter Newman. Biophilic architecture: a review of the rationale and outcomes[J]. AIMS Environmental Science, 2015, 2(4): 950-969. doi: 10.3934/environsci.2015.4.950

| [1] | Fromm E (1964) The Heart of Man. New York, USA: Harper and Row. |
| [2] | Wilson EO (1984) Biophilia. Massachusetts: Harvard University Press. |
| [3] | Kellert SR, Wilson EO (1993) The Biophilia Hypothesis. Washington, D.C.: Island Press. |
| [4] | Kellert SR, Heerwagen J, Mador M (2008) Biophilic design: the theory, science, and practice of bringing buildings to life. Hoboken, N.J.: Wiley. |
| [5] | Kellert SR (2008) Dimensions, elements, and attributes of biophilic design. In: Kellert SR, Heerwagen J, Mador ML, editors. Biophilic Design. New Jersey, US: John Wiley and Sons. pp. 3-19. |
| [6] | Salingaros NA, Masden KG (2008) Neuroscience, the natural environment, and building design. In: Kellert SR, Heerwagen JH, Mador ML, editors. Biophilic Design: The theory, science, and practice of bringing buildings to life. New Jersey: John Wiley & Sons. pp. 59-83. |
| [7] |
Miller JR (2005) Biodiversity conservation and the extinction of experience. Trend Ecol Evol 20: 430-434. doi: 10.1016/j.tree.2005.05.013

|
| [8] | Orr D (1993) Chapter 14: Love It or Lose It: The Coming Biophilia Revolution. In: Kellert SR, Wilson EO, editors. The Biophilia Hypothesis. Washington, DC, USA: Island Press. pp. 415-440. |
| [9] |
Newman P, Matan A (2012) Human Health and Human Mobility. Curr Opin Env Sust 4: 420-426. doi: 10.1016/j.cosust.2012.08.005
|
| [10] | Beatley T (2011) Biophilic Cities: Integrating Nature into Urban Design and Planning. Washington DC, USA: Island Press. |
| [11] | Nesse RM, Williams GC (1995) Evolution and healing: the new science of Darwinian medicine London: Weidenfeld and Nicolson. |
| [12] | Burns G (2005) Chapter 16: Naturally happy, naturally healthy: The role of the environment in wellbeing. In: Huppert FA, Baylis N, Keverne B, editors. The Science of Wellbeing. Oxford, UK: Oxford University Press. pp. 405-434. |
| [13] | Alexander C (1977) A pattern language: towns, buildings, construction/Christopher Alexander, Sara Ishikawa, Murray Silverstein; Ishikawa S, Silverstein M, editors. New York: Oxford University Press. |
| [14] | Appleton J (1975) The experience of landscape. London: Wiley. |
| [15] | Heerwagen J, Gregory B (2008) Biophilia and sensory aesthetics. In: Kellert SR, Heerwagen JH, Mador ML, editors. Biophilic Design: The theory, science, and practice of bringing buildings to life. New Jersey: John Wiley & Sons. pp. 227-241. |
| [16] | Wilson EO (2008) The nature of human nature. In: S.R. K, Heerwagen J, Mador ML, editors. Biophilic Design. New Jersey: John Wiley & Sons. pp. 21-25. |
| [17] | Kellert SR (2008) Biophilia. In: Sven EJ, Brian F, editors. Encyclopedia of Ecology. Oxford: Academic Press. pp. 462-466. |
| [18] | Browning WD, Ryan CO, Clancy JO (2014) 14 Patterns of biophilic design. New York: Terrapin Bright Green LLC. |
| [19] | Ryan CO, Browning WO, Clancy JO, et al. (2014) Biophillic design patterns: Emerging nature-based parameters for health and wellbeing in the built environment. ArchNet Int J Architect Res 8: 62-76. |
| [20] | Kellert SR, Calabrese EF, The Practice of Biophilic Design. 2015. Available from: http://www.biophilic-design.com/ |
| [21] |
Pert CB, Snyder SH (1973) Opiate Receptor: Demonstration in Nervous Tissue. Science 179: 1011-1014. doi: 10.1126/science.179.4077.1011
|
| [22] | Pert CB (1997) Molecules of emotion: why you feel the way you feel New York: Scribner. |
| [23] | Ulrich RS (1979) Visual landscapes and psychological wellbeing (National Parks). Landsc Res 4: 17-23. |
| [24] |
Ulrich RS (1984) View through a window may influence recovery from surgery. Science 224: 420-421. doi: 10.1126/science.6143402
|
| [25] | Kaplan R, Kaplan S (1989) The experience of nature: a psychological perspective; Kaplan S, editor. Cambridge: Cambridge University Press. |
| [26] |
Ulrich RS, Simons RF, Losito BD, et al. (1991) Stress recovery during exposure to natural and urban environments. J Environ Psychol 11: 201-230. doi: 10.1016/S0272-4944(05)80184-7
|
| [27] |
Kaplan S (1995) The restorative benefits of nature: Toward an integrative framework. J Environ Psychol 15: 169-182. doi: 10.1016/0272-4944(95)90001-2
|
| [28] |
Li Q, Otsuka T, Kobayashi M, et al. (2011) Acute effects of walking in forest environments on cardiovascular and metabolic parameters. Eur J Appl Physiol 111: 2845-2853. doi: 10.1007/s00421-011-1918-z
|
| [29] |
Berman MG, Kross E, Krpan KM, et al. (2012) Interacting with nature improves cognition and affect for individuals with depression. J Affect Disorders 140: 300-305. doi: 10.1016/j.jad.2012.03.012
|
| [30] |
Matsunaga K, Park BJ, Kobayashi H, et al. (2011) Physiologically relaxing effect of a hospital rooftop forest on older women requiring care. J Am Geriatr Soc 59: 2162-2163. doi: 10.1111/j.1532-5415.2011.03651.x
|
| [31] |
Park BJ, Tsunetsugu Y, Kasetani T, et al. (2010) The physiological effects of Shinrin-yoku (taking in the forest atmosphere or forest bathing): evidence from field experiments in 24 forests across Japan. Environ Health Prev Med 15: 18-26. doi: 10.1007/s12199-009-0086-9
|
| [32] |
Tyrväinen L, Ojala A, Korpela K, et al. (2014) The influence of urban green environments on stress relief measures: A field experiment. J Environ Psychol 38: 1-9. doi: 10.1016/j.jenvp.2013.12.005
|
| [33] |
Berman MG, Jonides J, Kaplan S (2008) The Cognitive Benefits of Interacting with Nature. Psychol Sci 19: 1207-1212. doi: 10.1111/j.1467-9280.2008.02225.x
|
| [34] |
Berto R (2005) Exposure to restorative environments helps restore attentional capacity. J Environ Psychol 25: 249-259. doi: 10.1016/j.jenvp.2005.07.001
|
| [35] | Ivarsson CT, Hagerhall CM (2008) The perceived restorativeness of gardens-Assessing the restorativeness of a mixed built and natural scene type. Urban For Urban Gree 7: 107-118. |
| [36] | Hartig T, Bringslimark T, Patil GG (2008) Chapter 9: Restorative Environmental Design: What, When, Where and for Whom? In: Kellert SR, Heerwagen JH, Mador ML, editors. Biophilic Design. Hoboken, USA: John Wiley and Sons, Inc. pp. 133-152. |
| [37] |
Nanda U, Pati D, Ghamari H, et al. (2013) Lessons from neuroscience: form follows function, emotions follow form. Intell Build Int 5: 61-78. doi: 10.1080/17508975.2013.807767
|
| [38] |
Kuo FE, Sullivan WC (2001) Environment and crime in the inner city-Does vegetation reduce crime? Environ Behav 33: 343-367. doi: 10.1177/00139160121973025
|
| [39] |
Kuo FE (2001) Coping with Poverty: impacts of environment and attention in the inner city. Environ Behav 33: 5-34. doi: 10.1177/00139160121972846
|
| [40] | Guéguen N, Stefan J (2014) "Green Altruism": Short Immersion in Natural Green Environments and Helping Behavior. Environ Behav 2014: 1-19. |
| [41] |
Raanaas RK, Evensen KH, Rich D, et al. (2011) Benefits of indoor plants on attention capacity in an office setting. J Environ Psychol 31: 99-105. doi: 10.1016/j.jenvp.2010.11.005
|
| [42] | Park SH, Mattson RH (2008) Effects of flowering and foliage plants in hospital rooms on patients recovering from abdominal surgery. Horttechnology 18: 563-568. |
| [43] |
Moore EO (1981) A prison environment's effect on health care service demands. J Environ Syst 11: 17-34. doi: 10.2190/KM50-WH2K-K2D1-DM69
|
| [44] |
Ikei H, Komatsu M, Song CR, et al. (2014) The physiological and psychological relaxing effects of viewing rose flowers in office workers. J Physiol Anthropol 33: 1-5. doi: 10.1186/1880-6805-33-1
|
| [45] |
Nieuwenhuis M, Knight C, Postmes T, et al. (2014) The Relative Benefits of Green Versus Lean Office Space: Three Field Experiments. J Exp Psychol Appl 20: 199-214. doi: 10.1037/xap0000024
|
| [46] | Hagerhall CM, Laike T, Taylor R, et al. Human eeg responses to exact and statistical fractal patterns; 22nd International Association People-Environment Studies (IAPS) 2012 Jun 24-29; Glasgow: UK. |
| [47] | Salingaros NA (2012) Fractal Art and Architecture Reduce Phsiological Stress. J Biourbanism 2: 11-28. |
| [48] |
Biederman I, Vessel EA (2006) Perceptual Pleasure and the Brain: A novel theory explains why the brain craves information and seeks it through the senses. Am Sci 94: 247-253. doi: 10.1511/2006.59.995
|
| [49] |
Ode Å, Hagerhall CM, Sang N (2010) Analysing Visual Landscape Complexity: Theory and Application. Landsc Res 35: 111-131. doi: 10.1080/01426390903414935
|
| [50] |
Stamps AE (2002) Fractals, skylines, nature and beauty. Landsc Urban Plan 60: 163-184. doi: 10.1016/S0169-2046(02)00054-3
|
| [51] |
Taylor RP (2006) Reduction of physiological stress using fractal art and architecture. Leonardo 39: 245-251. doi: 10.1162/leon.2006.39.3.245
|
| [52] | BiophilicCities Homepage. 2015. Available from: http://biophiliccities.org/ |
| [53] | Tan PY (2013) Vertical Garden City: Singapore. Singapore: Straits Times Press Pte Ltd. |
| [54] | Yok TP, Chiang K, Chan D, et al. (2009) Vertical Greenery for the Tropics; Chiang K, Tan A, editors. Singapore: National Parks Board and Building and Contruction Authority. |
| [55] | Anders RM, Walker JB. Green roof stormwater performance in a southeastern US climate; 9th Annual Green Roof AND Wall Conference 2011 Nov 30-Dec 3; Philadelphia, USA. |
| [56] |
Mentens J, Raes D, Hermy M (2006) Green roofs as a tool for solving the rainwater runoff problem in the urbanized 21st century? Landsc Urban Plan 77: 217-226. doi: 10.1016/j.landurbplan.2005.02.010
|
| [57] |
Schroll E, Lambrinos J, Righetti T, et al. (2011) The role of vegetation in regulating stormwater runoff from green roofs in a winter rainfall climate. Ecol Eng 37: 595-600. doi: 10.1016/j.ecoleng.2010.12.020
|
| [58] |
Gregoire BG, Clausen JC (2011) Effect of a modular extensive green roof on stormwater runoff and water quality. Ecol Eng 37: 963-969. doi: 10.1016/j.ecoleng.2011.02.004
|
| [59] |
Lee JY, Moon HJ, Kim TI, et al. (2013) Quantitative analysis on the urban flood mitigation effect by the extensive green roof system. Environ Pollut 181: 257-261. doi: 10.1016/j.envpol.2013.06.039
|
| [60] | Ostendorf M, Retzlaff W, Thompson K, et al. Storm water runoff from green retaining wall systems; Cities Alive: 9th Annual Green Roof and Wall Conference 2011 Nov 30-Dec 3; Philadelphia, USA. |
| [61] | Burrows RM, Corragio MA. Living Walls: Integration of water re-use systems. In: Cities GRfH, editor; Cities Alive: 9th Annual Green Roof and Wall Conference 2011 Nov 30-Dec 3; Philadelphia, USA. |
| [62] |
Burian SJ, Nix SJ, Pitt RE, et al. (2000) Urban Wastewater Management in the United States: Past, Present, and Future. J Urban Technol 7: 33-62. doi: 10.1080/713684134
|
| [63] |
Rowe DB (2011) Green roofs as a means of pollution abatement. Environ Pollut 159: 2100-2110. doi: 10.1016/j.envpol.2010.10.029
|
| [64] | Wang R, Eckelman M, Zimmerman J (2013) Consequential Environmental and Economic Life Cycle Assessment of Green and Gray Stormwater Infrastructures for Combined Sewer Systems. Environ Sci Technol 47: 11189-11198. |
| [65] | Anacostia Watershed Society Green Roofs 2015 Available at http://www.anacostiaws.org/green-roofs |
| [66] | Seidl M, Gromaire M-C, Saad M, et al. (2013) Effect of substrate depth and rain-event history on the pollutant abatement of green roofs. Environ Pollut 183: 195-203. |
| [67] | Akbari H (2002) Shade trees reduce building energy use and CO2 emissions from power plants. Environmental Pollution 116, Supplement 1: S119-S126. |
| [68] |
Leung DYC, Tsui JY, Chen F, et al. (2011) Effects of Urban Vegetation on Urban Air Quality. Landsc Res 36: 173-188. doi: 10.1080/01426397.2010.547570
|
| [69] |
Sheweka S, Magdy AN (2011) The Living walls as an Approach for a Healthy Urban Environment. Energy Procedia 6: 592-599. doi: 10.1016/j.egypro.2011.05.068
|
| [70] | Plains CO2 Reduction Partnership. CO2 and Storage in the Region. Available from: http://www.undeerc.org/PCOR/Region/ |
| [71] | Carey PA, Guide to Phytoremediation: A Symbiotic Relationship with Plants, Water & Living Architecture. 2013. Available from: http://www.greenroofs.com/content/Phytoremediation-A-Symbiotic-Relationship-with-Plants-Water-and-Living-Architecture.htm |
| [72] | Miyawaki A (1998) Restoration of urban green environments based on the theories of vegetation ecology. Ecol Eng 11: 157-165. |
| [73] |
Ottelé M, Perini K, Fraaij ALA, et al. (2011) Comparative life cycle analysis for green façades and living wall systems. Energy Build 43: 3419-3429. doi: 10.1016/j.enbuild.2011.09.010
|
| [74] |
Pugh TAM, MacKenzie AR, Whyatt JD, et al. (2012) Effectiveness of Green Infrastructure for Improvement of Air Quality in Urban Street Canyons. Environ Sci Technol 46: 7692-7699. doi: 10.1021/es300826w
|
| [75] | Ottelé M, van Bohemen HD, Fraaij ALA (2010) Quantifying the deposition of particulate matter on climber vegetation on living walls. Ecol Eng 36: 154-162. |
| [76] |
Wolverton B, McDonald R, Watkins E (1984) Foliage plants for removing indoor air pollutants from energy-efficient homes. Econ Bot 38: 224-228. doi: 10.1007/BF02858837
|
| [77] |
Pegas PN, Alves CA, Nunes T, et al. (2012) Could Houseplants Improve Indoor air Quality in Schools? J Toxicol Environ Health Part A 75: 1371-1380. doi: 10.1080/15287394.2012.721169
|
| [78] | Levin H, Can House Plants Solve Indoor Air Quality Problems? 2014. Available from: http://www.practicalasthma.net/pages/topics/aaplants.htm |
| [79] | CBD International, 2011-2020 United Nations Decade on Biodiversity. 2012. Available from: https://www.cbd.int/doc/press/2012/pr-2012-10-20-cop-11-en.pdf |
| [80] |
Kowarik I (2011) Novel urban ecosystems, biodiversity, and conservation. Environ Pollut 159: 1974-1983. doi: 10.1016/j.envpol.2011.02.022
|
| [81] |
Cook-Patton SC, Bauerle TL (2012) Potential benefits of plant diversity on vegetated roofs: A literature review. J Environ Manag 106: 85-92. doi: 10.1016/j.jenvman.2012.04.003
|
| [82] | Grant G (2006) Extensive Green Roofs in London. Urban Habitat 4: 51-65. |
| [83] | Brenneisan S (2006) Space for Urban Wildlife: Designing green Roofs as Habitats in Switzerland. Urban Habitat 4: 27-36. |
| [84] | Baumann N (2006) Ground-Nesting Birds on green Roofs in Switzerland: Preliminary Observations. Urban Habitat 4: 37-50. |
| [85] |
Madre F, Vergnes A, Machon N, et al. (2014) Green roofs as habitats for wild plant species in urban landscapes: First insights from a large-scale sampling. Landscape Urban Plan 122: 100-107. doi: 10.1016/j.landurbplan.2013.11.012
|
| [86] |
Newman P (2014) Biophilic urbanism: a case study on Singapore. Aust Plan 51: 47-65. doi: 10.1080/07293682.2013.790832
|
| [87] |
Wong NH, Kwang Tan AY, Chen Y, et al. (2010) Thermal evaluation of vertical greenery systems for building walls. Build Environ 45: 663-672. doi: 10.1016/j.buildenv.2009.08.005
|
| [88] |
Kontoleon KJ, Eumorfopoulou EA (2010) The effect of the orientation and proportion of a plant-covered wall layer on the thermal performance of a building zone. Build Environ 45: 1287-1303. doi: 10.1016/j.buildenv.2009.11.013
|
| [89] |
Sheweka SM, Mohamed NM (2012) Green Facades as a New Sustainable Approach Towards Climate Change. Energy Procedia 18: 507-520. doi: 10.1016/j.egypro.2012.05.062
|
| [90] |
Susca T, Gaffin SR, Dell'Osso GR (2011) Positive effects of vegetation: Urban heat island and green roofs. Environ Pollut 159: 2119-2126. doi: 10.1016/j.envpol.2011.03.007
|
| [91] |
Cheng CY, Cheung KKS, Chu LM (2010) Thermal performance of a vegetated cladding system on facade walls. Build Environ 45: 1779-1787. doi: 10.1016/j.buildenv.2010.02.005
|
| [92] |
Jaffal I, Ouldboukhitine S-E, Belarbi R (2012) A comprehensive study of the impact of green roofs on building energy performance. Renew Energ 43: 157-164. doi: 10.1016/j.renene.2011.12.004
|
| [93] |
Hongming H, Jim CY (2010) Simulation of thermodynamic transmission in green roof ecosystem. Ecol Model 221: 2949-2958. doi: 10.1016/j.ecolmodel.2010.09.002
|
| [94] |
Sproul J, Wan MP, Mandel BH, et al. (2014) Economic comparison of white, green and black flat roofs in the United States. Energy Build 71: 20-27. doi: 10.1016/j.enbuild.2013.11.058
|
| [95] | Browning B, Garvin C, Ryan C, et al., The Economics of Biophilia-Why Designing with Nature in Mind Makes Financial Sense. 2012. Available from: http://www.terrapinbrightgreen.com/report/economics-of-biophilia/ |
| [96] |
Heerwagen J (2000) Green buildings, organizational success and occupant productivity. Build Res Inf 28: 353-367. doi: 10.1080/096132100418500
|
| [97] | Erwine B, Heschong L (2000) Daylight: Healthy, wealthy & wise. Architect Light 15: 98. |
| [98] | Heschong L (2002) Daylighting and human performance. ASHRAE J 44: 65-67. |
| [99] | Flew T (2008) Not Yet the Internet Election: Online Media, Political Commentary and the 2007 Australian Federal Election. Media Int Aust Incorp Cult Policy 126: 5-13. |
| [100] |
Singh AMS, Syal M, Grady SC, et al. (2010) Effects of Green Buildings on Employee Health and Productivity. Am J Public Health 100: 1665-1668. doi: 10.2105/AJPH.2009.180687
|
| [101] |
Beauchemin KM, Hays P (1996) Sunny hospital rooms expedite recovery from severe and refractory depressions. J Affect Disorders 40: 49-51. doi: 10.1016/0165-0327(96)00040-7
|
| [102] |
Ulrich RS (2006) Essay: Evidence-based health-care architecture. Lancet 368: S38-S39. doi: 10.1016/S0140-6736(06)69921-2
|
| [103] |
Benedetti F, Colombo C, Barbini B, et al. (2001) Morning sunlight reduces length of hospitalization in bipolar depression. J Affect Disorders 62: 221-223. doi: 10.1016/S0165-0327(00)00149-X
|
| [104] | Ulrich RS. Health benefits of gardens in hospitals; Plants for People: International Exhibition Floriade 2002 April 6-October 20; Haarlemmerme, NETH. |
| [105] | Wolf Kl (2005) Trees in the small city retail business district: Comparing resident and visitor perceptions. J Forest 103: 390-395. |
| [106] |
Joye Y, Willems K, Brengman M, et al. (2010) The effects of urban retail greenery on consumer experience: Reviewing the evidence from a restorative perspective. Urban For Urban Gree 9: 57-64. doi: 10.1016/j.ufug.2009.10.001
|
| [107] |
Brengman M, Willems K, Joye Y (2012) The Impact of In Store Greenery on Customers. Psychol Mark 29: 807-821. doi: 10.1002/mar.20566
|
| [108] | Eichholtz P, Kok N, Quigley J (2010) Doing Well by Doing Good? Green Office Buildings. Am Econ Rev 100: 2492-2509. |
| [109] |
Benson ED, Hansen JL, Schwartz Jr AL, et al. (1998) Pricing Residential Amenities: The Value of a View. J Real Estate Financ Econ 16: 55-73. doi: 10.1023/A:1007785315925
|
| [110] | Newman P, Kenworthy J (2015) The End of Automobile Dependence: How Cities are Moving Beyond Car-Based Planning. New York, USA: Island Press. |
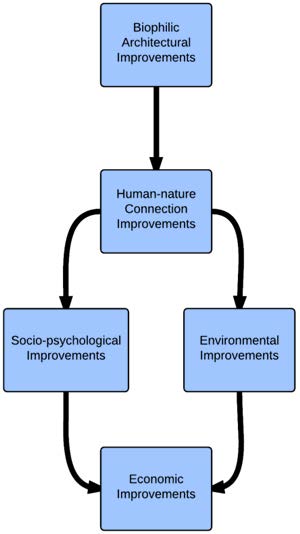
Figures(1) / Tables(1)

Jana Söderlund, Peter Newman. Biophilic architecture: a review of the rationale and outcomes[J]. AIMS Environmental Science, 2015, 2(4): 950-969. doi: 10.3934/environsci.2015.4.950







 DownLoad:
DownLoad: