Citation: Edward F. Durner. Plant architecture of Albion strawberry (Fragaria × ananassa Duch.) is not influenced by light source during conditioning[J]. AIMS Agriculture and Food, 2018, 3(3): 246-265. doi: 10.3934/agrfood.2018.3.246

| [1] | Durner E (2017) Long-day and nitrogen conditioning of 'Albion' strawberry (Fragaria × ananassa Duch.) enhances off-season field production. J Hortic Sci Biotechnol: 1–10. |
| [2] | Durner E (2017) Conditioning alleviates reduced yield of smaller plugs of 'Albion' strawberry (Fragaria × ananassa Duch.) in off-season plasticulture production. J Hortic Sci Biotechnol: 1–8. |
| [3] |
Delm TV, Melis P, Stoffels K, et al. (2013) Pre-harvest night-interruption on everbearing cultivars in out-of-soil strawberry cultivation in Belgium. Int J Fruit Sci 13: 217–226. doi: 10.1080/15538362.2012.698176

|
| [4] | Delm TV, Melis P, Stoffels K, et al. (2016) The effect of long-day treatment on runners and inflorescences on everbearing strawberry cultivar 'Capri'. Acta Hortic: 285–290. |
| [5] | Hamano M, Kato K, Honda K, et al. (2015) Promotion of flowering by photoperiod treatment in six strawberry (Fragaria × ananassa Duch.) cultivars with different everbearing patterns. J Pomol Hortic Sci 90: 157–163. |
| [6] | Heide OM, Stavang JA, Sonsteby AS (2015) Physiology and genetics of flowering in cultivated and wild strawberries – a review. J Pomol Hortic Sci 88: 1–18. |
| [7] |
Sonsteby A, Opstad N, Myrheim U, et al. (2009) Interaction of short day and timing of nitrogen fertilization on growth and flowering of 'Korona' strawberry (Fragaria × ananassa Duch.). Sci Hortic 123: 204–209. doi: 10.1016/j.scienta.2009.08.009
|
| [8] | Desmet EM, Verbraeken L, Baets W, et al. (2009) Optimisation of nitrogen fertilization prior to and during flowering process on performance of short day strawberry 'Elsanta'. Acta Hortic 842: 675–678. |
| [9] | Yamasaki A, Yano T, Lopezmedina J (2009) Effect of supplemental application of fertilizers on flower bud initiation and development of strawberry – possible role of nitrogen. Acta Hortic 842: 765–768. |
| [10] |
Sonsteby A, Opstad N, Heide O (2013) Environmental manipulation for establishing high yield potential of strawberry forcing plants. Sci Hortic 157: 65–73. doi: 10.1016/j.scienta.2013.04.014
|
| [11] | Durner E (2017) Programmed flowering of the F1 long-day strawberry cultivar 'Elan' with nitrogen and daylength manipulation. AIMS Agric Food 1: 4–19. |
| [12] |
Durner E (2016) Enhanced flowering of the F1 long-day strawberry cultivars 'Tarpan' and 'Gasana' with nitrogen and daylength management. AIMS Agric Food 2: 1–15. doi: 10.3934/agrfood.2017.1.1
|
| [13] |
Durner E (2017) Fall nitrogen enhances spring nitrogen enhanced flowering in the long day strawberry cultivar 'Elan'. AIMS Agric Food 2: 149–164. doi: 10.3934/agrfood.2017.2.149
|
| [14] |
Barthelemy D, Caraglio Y (2007) Plant architecture: A dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Ann Bot 99: 375–407. doi: 10.1093/aob/mcl260
|
| [15] | Savini G, Neri D (2004) Strawberry architectural model. Acta Hortic 649: 169–176. |
| [16] | Melis P (2013) Flower bud analysis of strawberry plants: predicting flower trusses and their spreading potential. Available from: http://www.hoogstraten.eu/congress/data/scientific/Predicting%20potential%20flower%20trusses%20and%20their%20spread%20through%20flower%20bud%20analysis%20of%20strawberry%20plants.pdf. |
| [17] | Gianluca S, Letouze A, Sabbadini C, et al. (2006) Evaluation of tray-plant quality in the propagation phase. Acta Hortic 74: 231–236. |
| [18] | Bosca JP, Neri D, Massetani F, et al. (2012) Relationship between plant architecture and fruit production of the short-day strawberry cultivar Gariguette. J Berry Res 2: 105–111. |
| [19] | Wobbrock J, Findlater L, Gergle D, et al. (2011) The aligned rank transform for nonparametric factorial analyses using only anova procedures. Sigchi Conf Hum Factors Comput Syst: 143–146. |
| [20] | Sonsteby AS, Heide O (2007) Quantitative long-day flowering response in the perpetual-flowering F1 strawberry cultivar 'Elan'. J Pomol Hortic Sci 82: 266–274. |
| [21] |
Sonsteby A, Heide O (2007) Long-day control of flowering in everbearing strawberries. J Hortic Sci Biotechnol 82: 875–884. doi: 10.1080/14620316.2007.11512321
|
| [22] | Lieten P (2002) Effect of nutrition prior to and during flower differentiation on phyllody and plant performance of short day strawberry 'Elsanta'. Acta Hortic 567: 345–348. |
| [23] |
Miere PL, Hadley P, Darby J, et al. (1996) The effect of temperature and photoperiod on the rate of flower initiation and the onset of dormancy in strawberry (Fragaria × ananassa Duch.). J Pomol Hortic Sci 71: 361–371. doi: 10.1080/14620316.1996.11515415
|
| [24] | Durner E (2016) Photoperiod and Temperature Conditioning of 'Sweet Charlie' Strawberry (Fragaria × ananassa Duch.) Plugs Enhances Off-Season Production. Sci Hortic 201: 184–189. |
| [25] | Furuya S, Yamashita M, Yamasaki A (1988) Effects of nitrogen content on the flower bud initiation induced by chilling under dark condition in strawberries. Japan, Kurume, Bull Natl Res Veg, Plants Tea, Series D. |
| [26] | Van den Muijzenberg E (1942) The influence of light and temperature on the periodic development of the strawberry and its significance in cultivation. Wageningen, the Netherlands, Ph.D. thesis, Laboratorium voor Tuinbouwplantenteelt, 160. |
| [27] |
Arney S (1953) The initiation, growth and emergence of leaf primordia in Fragaria. Ann Bot 17: 477–492. doi: 10.1093/oxfordjournals.aob.a083364
|
| [28] | Hytonen T, Palonen P, Mouhu K, et al. (2015) Crown branching and cropping potential in strawberry (Fragaria × ananassa Duch.) can be enhanced by daylength treatments. J Pomol Hortic Sci 79: 466–471. |
| [29] |
Durner E, Poling E (1987) Flower bud induction, initiation, differentiation and development in the 'Earliglow' strawberry. Sci Hortic 31: 61–69. doi: 10.1016/0304-4238(87)90107-5
|
| [30] |
Bowman G (2012) Strawberry production in forced and protected culture in Europe as a response to climate change. Can J Plant Sci 92: 1021–1036. doi: 10.4141/cjps2011-276
|
| [31] |
Yamasaki A (2013) Recent progress if strawberry year-round production technology in Japan. Jpn Agric Res Q 47: 37–42. doi: 10.6090/jarq.47.37
|
| [32] | Guttridge C (1985) Fragaria × ananassa, In: Halevy, A.H. (Ed.), Handbook of Flowering, vol 3., Boca Raton: CRC Press, 16–33. |
| [33] | Mochizuki T (1995) Past and present strawberry breeding programs in Japan. Adv Strawberry Res 14: 9–17. |
| [34] | Anderson H, Guttridge C (1982) Strawberry truss morphology and the fate of high-order flower buds. Crop Res 22: 105–122. |
| [35] | Fujimoto K, Kimura M (1970) Studies on flowering of strawberry III, Effect of nitrogen on flower bud differentiation and development. Japanese: Abstracts of the Japanese Society for Horticultural Science Spring Meeting, 174–175. |
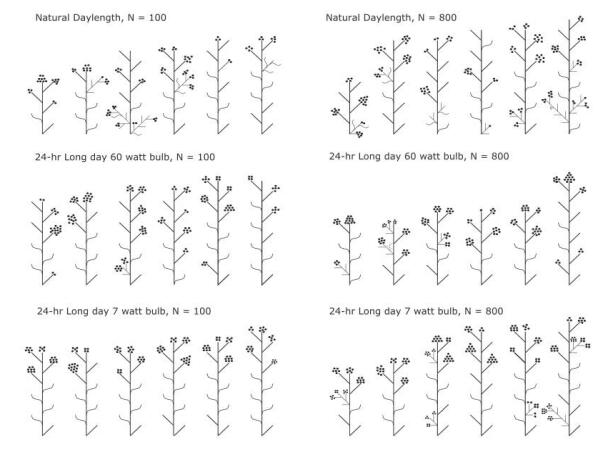
Figures(1) / Tables(10)

Edward F. Durner. Plant architecture of Albion strawberry (Fragaria × ananassa Duch.) is not influenced by light source during conditioning[J]. AIMS Agriculture and Food, 2018, 3(3): 246-265. doi: 10.3934/agrfood.2018.3.246







 DownLoad:
DownLoad: