Citation: Alfred Mitema, Naser Aliye Feto. Molecular and Vegetative Compatibility Groups Characterization of Aspergillus flavus Isolates from Kenya[J]. AIMS Microbiology, 2020, 6(3): 231-249. doi: 10.3934/microbiol.2020015

| [1] |
Reddy KRN, Reddy CS, Muralidharan K, et al. (2009) Potential of botanicals and biocontrol agents on growth and aflatoxin production by Aspergillus flavus infecting rice grains. Food Control 20: 173-178. doi: 10.1016/j.foodcont.2008.03.009

|
| [2] | Okoth S (2016) Improving the evidence base on aflatoxin contamination and exposure in Africa. CTA Work Pap 16/13. |
| [3] | Klich MA (2002) Indentification of common Aspergillus species. Centraalbureau voor Schimmelcultures . |
| [4] |
Pildain MB, Vaamonde G, Cabral D, et al. (2004) Analysis of population structure of Aspergillus flavus from peanut based on vegetative compatibility, geographic origin, mycotoxin and sclerotia production. Int J Food Microbiol 93: 31-40. doi: 10.1016/j.ijfoodmicro.2003.10.007
|
| [5] |
Varga J, Frisvad JC, Kocsubé S, et al. (2011) New and revisited species in Aspergillus section Nigri Extrolite analysis. Stud Mycol 69: 1-17. doi: 10.3114/sim.2011.69.01
|
| [6] |
Leslie JF (1993) Fungal Vegetative Compatibility. Annu Rev Phytopathol 31: 127-50. doi: 10.1146/annurev.py.31.090193.001015
|
| [7] |
Cotty PJ (1989) Virulence and cultural characteristics of two Aspergillus flavus strains pathogenic on cotton. Phytopathology 79: 808-814. doi: 10.1094/Phyto-79-808
|
| [8] |
Xiang Q, Glass NL (2004) The control of mating type heterokaryon incompatibility by vib-1, a locus involved in het-c heterokaryon incompatibility in Neurospora crassa. Fungal Genet Biol 41: 1063-76. doi: 10.1016/j.fgb.2004.07.006
|
| [9] |
Horn BW, Dorner JW (2009) Effect of nontoxigenic Aspergillus flavus and A. parasiticus on aflatoxin contamination of wounded peanut seeds inoculated with agricultural soil containing natural fungal populations. Biocontrol Sci Technol 9: 249-262. doi: 10.1080/09583150802696541
|
| [10] |
Grubisha LC, Cotty PJ (2010) Genetic isolation among sympatric vegetative compatibility groups of the aflatoxin-producing fungus Aspergillus flavus. Mol Ecol 19: 269-80. doi: 10.1111/j.1365-294X.2009.04467.x
|
| [11] |
Atehnkeng J, Donner M, Peter S, et al. (2016) Environmental distribution and genetic diversity of vegetative compatibility groups determine biocontrol strategies to mitigate aflatoxin contamination of maize by Aspergillus flavus. Microb Biotechnol 9: 75-88. doi: 10.1111/1751-7915.12324
|
| [12] | Mitema AO (2018) Molecular and Bio-analytical Characterisation as a means to understand Genetic diversity within Kenyan Aspergillus flavus strains.South Africa: University of Cape Town. |
| [13] |
Mitema A, Okoth S, Rafudeen MS, et al. (2018) Vegetative compatibility and phenotypic characterization as a means of determining genetic diversity of Aspergillus flavus isolates. Fungal Biol 122. doi: 10.1016/j.funbio.2017.11.005
|
| [14] |
Bayman P, Cotty PJ (1993) Genetic diversity in Aspergillus flavus: association with aflatoxin production and morphology. Can J Bot 71: 23-31. doi: 10.1139/b93-003
|
| [15] |
Mehl HL, Cotty PJ (2011) Influence of the Host Contact Sequence on the Outcome of Competition among Aspergillus flavus Isolates during Host Tissue Invasion. Appl Environ Microbiol 77: 1691-1697. doi: 10.1128/AEM.02240-10
|
| [16] |
Atehnkeng J, Ojiambo PS, Ikotun T, et al. (2008) Evaluation of atoxigenic isolates of Aspergillus flavus as potential biocontrol agents for aflatoxin in maize. Food Addit Contam Part A 25: 1264-71. doi: 10.1080/02652030802112635
|
| [17] |
Asao T (1996) Single-strand conformation polymorphism analysis of PCR-amplified ribosomal DNA Internal Transcribed Spacers to differentiate species of Aspergillus section Flavi. Appl Environ Microbiol 62: 2947-2952. doi: 10.1128/AEM.62.8.2947-2952.1996
|
| [18] |
Balajee SA, Houbraken J, Verweij PE, et al. (2007) Aspergillus species identification in the clinical setting. Stud Mycol 59: 39-46. doi: 10.3114/sim.2007.59.05
|
| [19] |
Clark K, Karsch-Mizrachi I, Lipman DJ, et al. (2016) GenBank. Nucleic Acids Res 44: D67-72. doi: 10.1093/nar/gkv1276
|
| [20] | Nilsson RH, Kristiansson E, Ryberg M, et al. (2008) Intraspecific ITS variability in the Kingdom Fungi as expressed in the international sequence databases and its implications for molecular species identification. Evol Bioinforma 2008: 193-201. |
| [21] |
Schoch CL, Seifert KA, Huhndorf S, et al. (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. PNAS 109: 6241-6. doi: 10.1073/pnas.1117018109
|
| [22] |
Frisvad JC, Skouboe P, Samson RA, et al. (2005) Taxonomic comparison of three different groups of aflatoxin producers and a new efficient producer of aflatoxin B<inf>1</inf>, sterigmatocystin and 3-O-methylsterigmatocystin, Aspergillus rambellii sp. nov. Syst Appl Microbiol 28: 442-453. doi: 10.1016/j.syapm.2005.02.012
|
| [23] |
Pildain MB, Frisvad JC, Vaamonde G, et al. (2008) Two novel aflatoxin-producing Aspergillus species from Argentinean peanuts. Int J Syst Evol Microbiol 58: 725-35. doi: 10.1099/ijs.0.65123-0
|
| [24] | FAO (2004) Mycotoxin regulations in 2003 and current developments. Worldw Regul mycotoxins food Feed 2003 9–28. |
| [25] |
Lewis LMO, Henry Njapau, Helen Schurz-Rogers, et al. (2005) Aflatoxin Contamination of Commercial Maize Products during an Outbreak of Acute Aflatoxicosis in Eastern and Central Kenya. Environ Health Perspect 113: 1763-7. doi: 10.1289/ehp.7998
|
| [26] |
Clark R, Steel D (2007) Sampling within households in household surveys. J R Stat Soc Ser A 170: 63-82. doi: 10.1111/j.1467-985X.2006.00434.x
|
| [27] |
Corsi D, Neuman M, Finlay J, et al. (2012) Demographic and health surveys: a profile. Int J Epidemiol 41: 1602-13. doi: 10.1093/ije/dys184
|
| [28] |
Okoth S, Nyongesa B, Ayugi V, et al. (2012) Toxigenic Potential of Aspergillus species Occurring on Maize Kernels from Two Agro-Ecological Zones in Kenya. Toxins (Basel) 4: 991-1007. doi: 10.3390/toxins4110991
|
| [29] |
Bayman P, Cotty PJ (1991) Vegetative compatibility and genetic diversity in the Aspergillus flavus population of a single field. Can J Bot 69: 1707-1711. doi: 10.1139/b91-216
|
| [30] |
Cove DJ (1976) Chlorate toxicity in Aspergillus nidulans: the selection and characterisation of chlorate resistant mutants. Heredity (Edinb) 36: 191-203. doi: 10.1038/hdy.1976.24
|
| [31] |
Untergasser A, Cutcutache I, Koressaar T, et al. (2012) Primer3-new capabilities and interfaces. Nucleic Acids Res 40: 1-12. doi: 10.1093/nar/gks596
|
| [32] |
Tamura K, Stecher G, Peterson D, et al. (2013) MEGA6: Molecular Evolutionary Genetics Analysis Version 6. 0. Mol Biol Evol 30: 2725-2729. doi: 10.1093/molbev/mst197
|
| [33] | Tamura K (1992) Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol Biol Evol 9: 678-87. |
| [34] |
Felsenstein J (1985) Phylogenies and the Comparative Method. Am Nat 125: 1-15. doi: 10.1086/284325
|
| [35] |
Samson RA, Visagie CM, Houbraken J, et al. (2014) Phylogeny, identification and nomenclature of the genus Aspergillus. Stud Mycol 78: 141-73. doi: 10.1016/j.simyco.2014.07.004
|
| [36] |
Moore GG, Horn BW (2009) Sexual Reproduction in Aspergillus flavus. Mycologia 101: 423-429. doi: 10.3852/09-011
|
| [37] | Papa (1986) Heterokaryon Incompatibility in Aspergillus flavus. Mycologia 78: 98-101. |
| [38] | Barros G, Chiotta ML, Torres A, et al. (2006) Genetic diversity in Aspergillus parasiticus population from the peanut agroecosystem in Argentina. Lett Appl Microbiol 42: 560-566. |
| [39] |
Horn BW, Greene RL (1995) Vegetative compatibility within populations of Aspergillus flavus, A. parasiticus, and A. tamarii from a peanut field. Mycologia 87: 324-332. doi: 10.1080/00275514.1995.12026537
|
| [40] |
Atehnkeng J, Ojiambo PS, Ikotun T, et al. (2008) Evaluation of atoxigenic isolates of Aspergillus flavus as potential biocontrol agents for aflatoxin in maize. Food Addit Contam 25: 1264-1271. doi: 10.1080/02652030802112635
|
| [41] |
Bock CH, Mackey B, Cotty PJ, et al. (2004) Population dynamics of Aspergillus flavus in the air of an intensively cultivated region of south-west Arizona. Plant Pathol 53: 422-433. doi: 10.1111/j.0032-0862.2004.01015.x
|
| [42] | Cotty J (2006) Biocompetitive exclusion of toxigenic fungi. Mycotoxin Factb 179-197. |
| [43] |
Gonçalves JS, Ferracin LM, Vieira MLC, et al. (2012) Molecular analysis of Aspergillus section Flavi isolated from Brazil nuts. World J Microbiol Biotechnol 28: 1817-1825. doi: 10.1007/s11274-011-0956-3
|
| [44] |
Gonçalves SS, Cano JF, Stchigel AM, et al. (2012) Molecular phylogeny and phenotypic variability of clinical and environmental strains of Aspergillus flavus. Fungal Biol 116: 1146-1155. doi: 10.1016/j.funbio.2012.08.006
|
| [45] |
Geiser DM, Dorner JW, Horn BW, et al. (2000) The Phylogenetics of Mycotoxin and Sclerotium Production in Aspergillus flavus and Aspergillus oryzae. Fungal Genet Biol 31: 169-179. doi: 10.1006/fgbi.2000.1215
|
| [46] |
Chang P, Ehrlich KC, Hua ST, et al. (2006) Cladal relatedness among Aspergillus oryzae isolates and Aspergillus flavus S and L morphotype isolates. Int J Food Microbiol 108: 172-177. doi: 10.1016/j.ijfoodmicro.2005.11.008
|
 microbiol-06-03-015-s001.pdf microbiol-06-03-015-s001.pdf |
 |
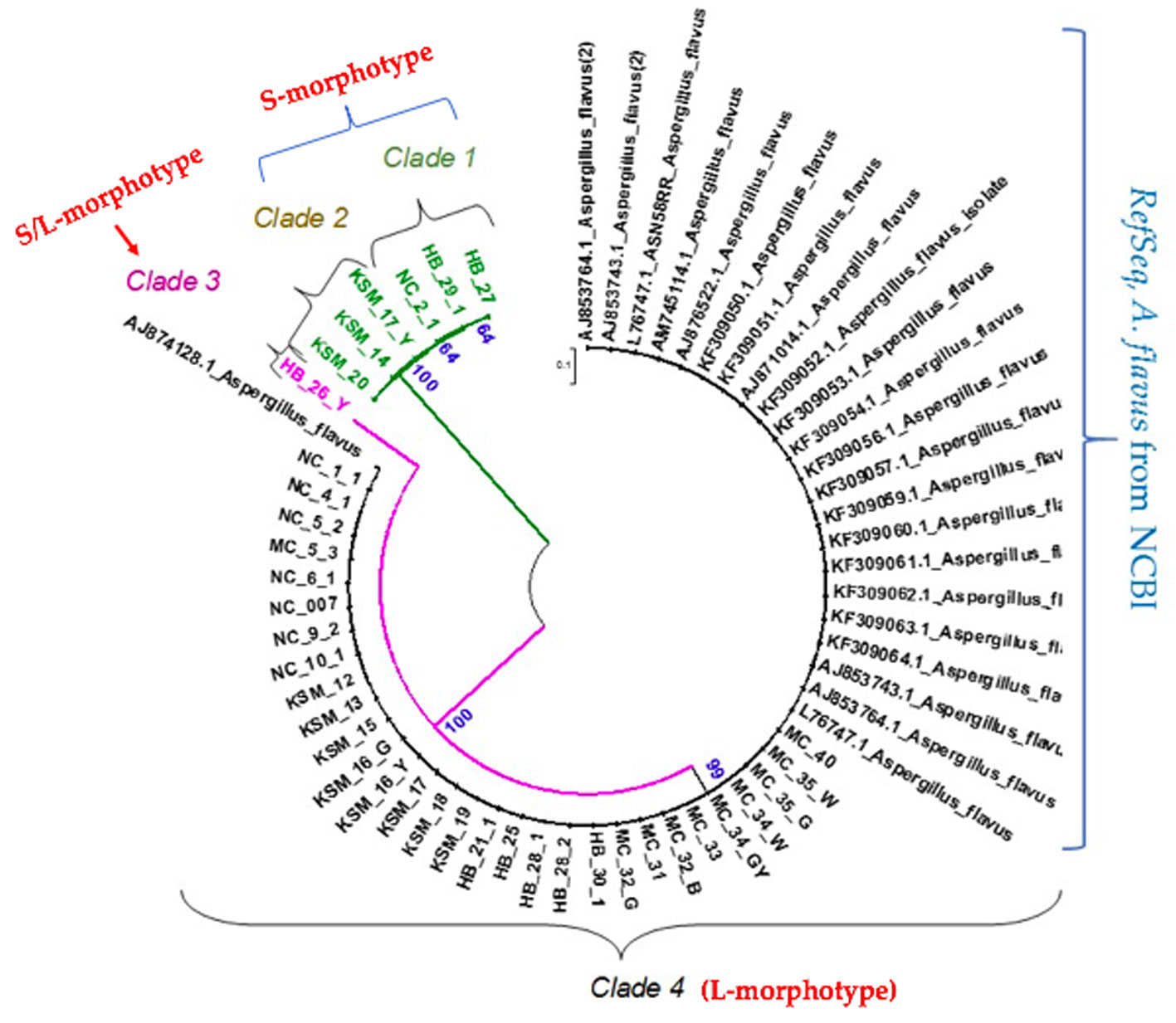
Figures(5) / Tables(2)

Alfred Mitema, Naser Aliye Feto. Molecular and Vegetative Compatibility Groups Characterization of Aspergillus flavus Isolates from Kenya[J]. AIMS Microbiology, 2020, 6(3): 231-249. doi: 10.3934/microbiol.2020015







 DownLoad:
DownLoad: