Citation: Christopher D. Chambers, Eva Feredoes, Suresh D. Muthukumaraswamy, Peter J. Etchells. Instead of “playing the game” it is time to change the rules: Registered Reports at AIMS Neuroscience and beyond[J]. AIMS Neuroscience, 2014, 1(1): 4-17. doi: 10.3934/Neuroscience.2014.1.4

| [1] |
Ioannidis JPA. (2005) Why Most Published Research Findings Are False. PLoS Med 2: e124. doi: 10.1371/journal.pmed.0020124

|
| [2] |
John LK, Loewenstein G, Prelec D. (2012) Measuring the prevalence of questionable research practices with incentives for truth telling. Psychol Sci 23: 524-532. doi: 10.1177/0956797611430953
|
| [3] | Simmons JP, Nelson LD, Simonsohn U. (2011) False-positive psychology: Undisclosed flexibility in data collection and analysis allows presenting anything as significant. Psychol Sci 22: 359-366. |
| [4] |
Kerr NL. (1998) HARKing: hypothesizing after the results are known. Pers Soc Psychol Rev 2: 196-217. doi: 10.1207/s15327957pspr0203_4
|
| [5] |
Makel MC, Plucker JA, Hegarty B. (2012) Replications in Psychology Research: How Often Do They Really Occur? Perspect Psychol Sci 7: 537-542. doi: 10.1177/1745691612460688
|
| [6] |
Faneli D. (2010) “Positive” Results Increase Down the Hierarchy of the Sciences. PLos One 5: e10068. doi: 10.1371/journal.pone.0010068
|
| [7] |
Button KS, Ioannidis JP, Mokrysz C, Nosek BA, Flint J, et al. (2013) Power failure: why small sample size undermines the reliability of neuroscience. Nat Rev Neurosci 14: 365-376. doi: 10.1038/nrn3475
|
| [8] |
Wicherts JM, Bakker M, Molenaar D. (2011) Willingness to share research data is related to the strength of the evidence and the quality of reporting of statistical results. PLoS One 6: e26828. doi: 10.1371/journal.pone.0026828
|
| [9] |
Cohen J. (1962) The statistical power of abnormal-social psychological research: a review. J Abnorm Soc Psychol 65: 145-153. doi: 10.1037/h0045186
|
| [10] | Sterling TD. (1959) Publication Decisions and their Possible Effects on Inferences Drawn from Tests of Significance—or Vice Versa. J Am Stat Assoc 54: 30-34. |
| [11] |
de Groot AD. (2014) The meaning of "significance" for different types of research [translated and annotated by Eric-Jan Wagenmakers, Denny Borsboom, Josine Verhagen, Rogier Kievit, Marjan Bakker, Angelique Cramer, Dora Matzke, Don Mellenbergh, and Han L. J. van der Maas]. Acta Psychol (Amst) 148: 188-194. doi: 10.1016/j.actpsy.2014.02.001
|
| [12] | Nosek BA, Spies JR, Motyl M. (2012) Scientific Utopia : II. Restructuring Incentives and Practices to Promote Truth Over Publishability. Perspect Psychol Sci 7: 615-631. |
| [13] |
Ioannidis JPA. (2012) Why Science Is Not Necessarily Self-Correcting. Perspect Psychol Sci 7: 645-654. doi: 10.1177/1745691612464056
|
| [14] |
Chambers CD. (2013) Registered reports: a new publishing initiative at Cortex. Cortex 49: 609-610. doi: 10.1016/j.cortex.2012.12.016
|
| [15] |
Wolfe J. (2013) Registered Reports and Replications in Attention, Perception, & Psychophysics. Atten Percept Psycho 75: 781-783. doi: 10.3758/s13414-013-0502-5
|
| [16] |
Stahl C. (2014) Experimental psychology: toward reproducible research. Exp Psychol 61: 1-2. doi: 10.1027/1618-3169/a000257
|
| [17] |
Munafo MR, Strain E. (2014) Registered Reports: A new submission format at Drug and Alcohol Dependence. Drug Alcohol Depend 137: 1-2. doi: 10.1016/j.drugalcdep.2014.02.699
|
| [18] | Nosek BA, Lakens D. (in press) Registered reports: A method to increase the credibility of published results. Soc Psychol. |
| [19] | Chambers CD, Munafo MR. (2013) Trust in science would be improved by study pre-registration. The Guardian:http://www. theguardian. com/science/blog/2013/jun/2005/trust-in-science-study-pre-registration. |
| [20] | Scott SK. (2013) Will pre-registration of studies be good for psychology? : https://sites. google. com/site/speechskscott/SpeakingOut/willpre-registrationofstudiesbegoodforpsychology. |
| [21] | Scott SK. (2013) Pre-registration would put science in chains. Times Higher Education: http://www. timeshighereducation. co. uk/comment/opinion/science-in-chains/2005954. article. |
| [22] |
Rouder J, Speckman P, Sun D, Morey R, Iverson G. (2009) Bayesian t tests for accepting and rejecting the null hypothesis. Psychon Bull Rev 16: 225-237. doi: 10.3758/PBR.16.2.225
|
| [23] |
Wagenmakers EJ. (2007) A practical solution to the pervasive problems of p values. Psychon Bull Rev 14: 779-804. doi: 10.3758/BF03194105
|
| [24] |
Dienes Z. (2011) Bayesian Versus Orthodox Statistics: Which Side Are You On? Perspect Psychol Sci 6: 274-290. doi: 10.1177/1745691611406920
|
| [25] |
Mathieu S, Chan AW, Ravaud P. (2013) Use of trial register information during the peer review process. PLoS One 8: e59910. doi: 10.1371/journal.pone.0059910
|
| [26] | Gelman A, Loken E. (2014) The garden of forking paths: Why multiple comparisons can be a problem, even when there is no fishing expedition" or "p-hacking" and the research hypothesis was posited ahead of time. Unpublished manuscript: http://www. stat. columbia. edu/~gelman/research/unpublished/p_hacking. pdf. |
| [27] |
Strube MJ. (2006) SNOOP:a program for demonstrating the consequences of premature and repeated null hypothesis testing. Behav Res Methods 38: 24-27. doi: 10.3758/BF03192746
|
| [28] |
Fiedler K, Kutzner F, Krueger JI. (2012) The Long Way From α-Error Control to Validity Proper: Problems With a Short-Sighted False-Positive Debate. Perspect Psychol Sci 7: 661-669. doi: 10.1177/1745691612462587
|
| [29] |
Whelan R, Conrod PJ, Poline JB, Lourdusamy A, Banaschewski T, et al. (2012) Adolescent impulsivity phenotypes characterized by distinct brain networks. Nat Neurosci 15: 920-925. doi: 10.1038/nn.3092
|
| [30] | Brembs B, Button K, Munafo M. (2013) Deep impact: unintended consequences of journal rank. Front Hum Neurosci 7: 291. |
| [31] | Nelson LD. (2014) Preregistration: Not just for the Empiro-zealots. http://datacoladaorg/2014/01/07/12-preregistration-not-just-for-the-empiro-zealots/. |
| [32] |
World Medical A (2013) World medical association declaration of helsinki: Ethical principles for medical research involving human subjects. JAMA 310: 2191-2194. doi: 10.1001/jama.2013.281053
|
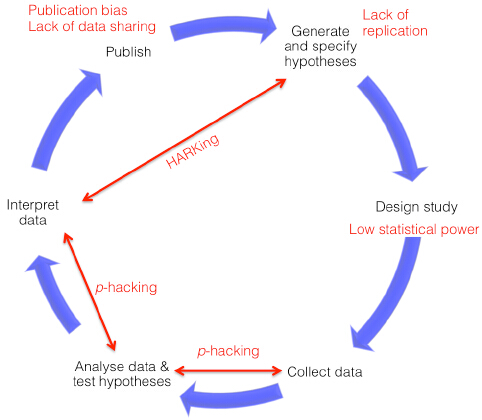

Figures(2)

Christopher D. Chambers, Eva Feredoes, Suresh D. Muthukumaraswamy, Peter J. Etchells. Instead of “playing the game” it is time to change the rules: Registered Reports at AIMS Neuroscience and beyond[J]. AIMS Neuroscience, 2014, 1(1): 4-17. doi: 10.3934/Neuroscience.2014.1.4







 DownLoad:
DownLoad: