Citation: David Petillo, Stephen Orey, Aik Choon Tan, Lars Forsgren, Sok Kean Khoo. Parkinsons Disease-related Circulating microRNA Biomarkers——a Validation Study[J]. AIMS Medical Science, 2015, 2(1): 7-14. doi: 10.3934/medsci.2015.1.7

| [1] |
Margis R, Rieder CR (2011) Identification of blood microRNAs associated to Parkinsonis disease. J biotechnol 152: 96-101. doi: 10.1016/j.jbiotec.2011.01.023

|
| [2] |
Martins M, Rosa A, Guedes LC, et al. (2011) Convergence of miRNA expression profiling, alpha-synuclein interacton and GWAS in Parkinson's disease. PloS one 6: e25443. doi: 10.1371/journal.pone.0025443
|
| [3] | Khoo SK, Petillo D, Kang UJ, et al. (2012) Plasma-based circulating microRNA biomarkers for Parkinson's disease. J Parkinson Dis 2: 321-331. |
| [4] |
Cardo LF, Coto E, de Mena L, et al. (2013) Profile of microRNAs in the plasma of Parkinson's disease patients and healthy controls. J Neurol 260: 1420-1422. doi: 10.1007/s00415-013-6900-8
|
| [5] | Soreq L, Salomonis N, Bronstein M, et al. (2013) Small RNA sequencing-microarray analyses in Parkinson leukocytes reveal deep brain stimulation-induced splicing changes that classify brain region transcriptomes. Front Mol Neurosci 6: 10. |
| [6] | Vallelunga A, Ragusa M, Di Mauro S, et al. (2014) Identification of circulating microRNAs for the differential diagnosis of Parkinson's disease and Multiple System Atrophy. Front Cell Neurosci8: 156. |
| [7] |
Botta-Orfila T, Morato X, Compta Y, et al. (2014) Identification of blood serum micro-RNAs associated with idiopathic and LRRK2 Parkinson's disease. J Neurosci Res 92: 1071-1077. doi: 10.1002/jnr.23377
|
| [8] |
Burgos K, Malenica I, Metpally R, et al. (2014) Profiles of extracellular miRNA in cerebrospinal fluid and serum from patients with Alzheimer's and Parkinson's diseases correlate with disease status and features of pathology. PLoS One 9: e94839. doi: 10.1371/journal.pone.0094839
|
| [9] |
Khoo SK, Neuman LA, Forsgren L, et al. (2013) Could miRNA expression changes be a reliable clinical biomarker for Parkinson's disease? Neurodegener Dis Manag 3: 455-465. doi: 10.2217/nmt.13.53
|
| [10] |
Kroh EM, Parkin RK, Mitchell PS, et al. (2010) Analysis of circulating microRNA biomarkers in plasma and serum using quantitative reverse transcription-PCR (qRT-PCR). Methods 50: 298-301. doi: 10.1016/j.ymeth.2010.01.032
|
| [11] |
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25: 402-408. doi: 10.1006/meth.2001.1262
|
| [12] |
Warnock DG, Peck CC (2010) A roadmap for biomarker qualification. Nat Biotechnol 28:444-445. doi: 10.1038/nbt0510-444
|
| [13] |
Tan AC, Naiman DQ, Xu L, et al. (2005) Simple decision rules for classifying human cancers from gene expression profiles. Bioinformatics 21: 3896-3904. doi: 10.1093/bioinformatics/bti631
|
| [14] |
Paulovich AG, Whiteaker JR, Hoofnagle AN, et al. (2008) The interface between biomarker discovery and clinical validation: The tar pit of the protein biomarker pipeline. Proteomics Clin Appl 2: 1386-1402. doi: 10.1002/prca.200780174
|
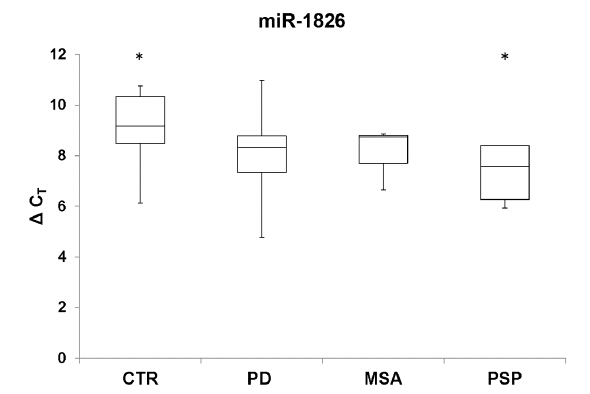
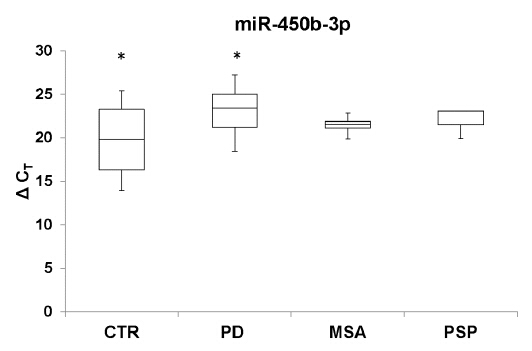
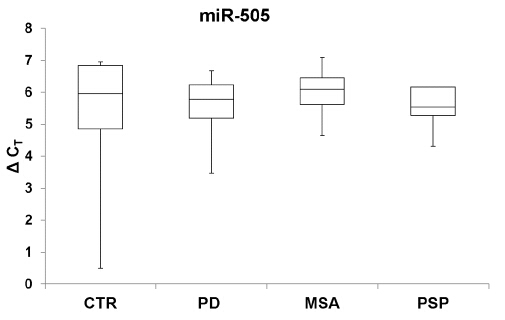
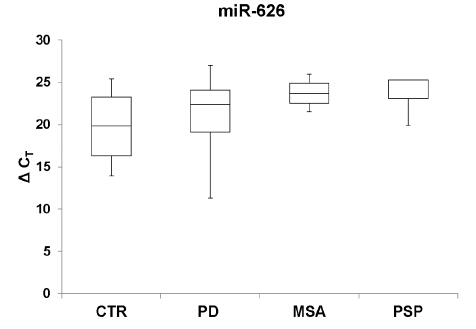
Figures(4) / Tables(2)

David Petillo, Stephen Orey, Aik Choon Tan, Lars Forsgren, Sok Kean Khoo. Parkinsons Disease-related Circulating microRNA Biomarkers——a Validation Study[J]. AIMS Medical Science, 2015, 2(1): 7-14. doi: 10.3934/medsci.2015.1.7







 DownLoad:
DownLoad: