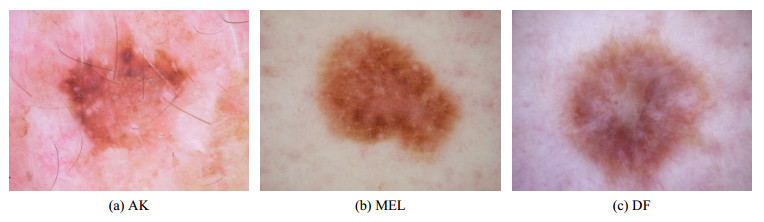
Methods based on deep learning have shown good advantages in skin lesion recognition. However, the diversity of lesion shapes and the influence of noise disturbances such as hair, bubbles, and markers leads to large intra-class differences and small inter-class similarities, which existing methods have not yet effectively resolved. In addition, most existing methods enhance the performance of skin lesion recognition by improving deep learning models without considering the guidance of medical knowledge of skin lesions. In this paper, we innovatively construct feature associations between different lesions using medical knowledge, and design a medical domain knowledge loss function (MDKLoss) based on these associations. By expanding the gap between samples of various lesion categories, MDKLoss enhances the capacity of deep learning models to differentiate between different lesions and consequently boosts classification performance. Extensive experiments on ISIC2018 and ISIC2019 datasets show that the proposed method achieves a maximum of 91.6% and 87.6% accuracy. Furthermore, compared with existing state-of-the-art loss functions, the proposed method demonstrates its effectiveness, universality, and superiority.
Citation: Li Zhang, Xiangling Xiao, Ju Wen, Huihui Li. MDKLoss: Medicine domain knowledge loss for skin lesion recognition[J]. Mathematical Biosciences and Engineering, 2024, 21(2): 2671-2690. doi: 10.3934/mbe.2024118
Methods based on deep learning have shown good advantages in skin lesion recognition. However, the diversity of lesion shapes and the influence of noise disturbances such as hair, bubbles, and markers leads to large intra-class differences and small inter-class similarities, which existing methods have not yet effectively resolved. In addition, most existing methods enhance the performance of skin lesion recognition by improving deep learning models without considering the guidance of medical knowledge of skin lesions. In this paper, we innovatively construct feature associations between different lesions using medical knowledge, and design a medical domain knowledge loss function (MDKLoss) based on these associations. By expanding the gap between samples of various lesion categories, MDKLoss enhances the capacity of deep learning models to differentiate between different lesions and consequently boosts classification performance. Extensive experiments on ISIC2018 and ISIC2019 datasets show that the proposed method achieves a maximum of 91.6% and 87.6% accuracy. Furthermore, compared with existing state-of-the-art loss functions, the proposed method demonstrates its effectiveness, universality, and superiority.

| [1] |
K. Doi, Computer-aided diagnosis in medical imaging: historical review, current status and future potential, Comput. Med. Imaging Graphics, 31 (2007), 198–211. https://doi.org/10.1016/j.compmedimag.2007.02.002 doi: 10.1016/j.compmedimag.2007.02.002

|
| [2] |
M. Ahammed, M. A. Mamun, M. S. Uddin, A machine learning approach for skin disease detection and classification using image segmentation, Healthcare Anal., 2 (2022), 100122. https://doi.org/10.1016/j.health.2022.100122 doi: 10.1016/j.health.2022.100122
|
| [3] |
J. Zhang, Y. Xia, Y. Xie, M. Fulham, D. D. Feng, Classification of medical images in the biomedical literature by jointly using deep and handcrafted visual features, IEEE J. Biomed. Health. Inf., 22 (2017), 1521–1530. https://doi.org/10.1109/JBHI.2017.2775662 doi: 10.1109/JBHI.2017.2775662
|
| [4] |
P. I. R. Jenifer, S. Kannan, Deep learning with optimal hierarchical spiking neural network for medical image classification, Comput. Syst. Sci. Eng., 44 (2023), 1081–1097. https://doi.org/10.32604/csse.2023.026128 doi: 10.32604/csse.2023.026128
|
| [5] |
J. Dominic, N. Bhaskhar, A. D. Desai, A. Schmidt, E. Rubin, B. Gunel, et al., Improving data-efficiency and robustness of medical imaging segmentation using inpainting-based self-supervised learning, Bioengineering, 10 (2023), 207. https://doi.org/10.3390/bioengineering10020207 doi: 10.3390/bioengineering10020207
|
| [6] |
L. Tan, H. Wu, J. Xia, Y. Liang, J. Zhu, Skin lesion recognition via global-local attention and dual-branch input network, Eng. Appl. Artif. Intell., 127 (2023), 107385. https://doi.org/10.1016/j.engappai.2023.107385 doi: 10.1016/j.engappai.2023.107385
|
| [7] |
F. Bozkurt, Skin lesion classification on dermatoscopic images using effective data augmentation and pre-trained deep learning approach, Multimedia Tools Appl., 82 (2023), 18985–19003. https://doi.org/10.1007/s11042-022-14095-1 doi: 10.1007/s11042-022-14095-1
|
| [8] |
X. Feng, T. Wang, X. Yang, M. Zhang, W. Guo, W. Wang, Convwin-unet: unet-like hierarchical vision transformer combined with convolution for medical image segmentation, Math. Biosci. Eng., 20 (2023), 128–144. https://doi.org/10.3934/mbe.2023007 doi: 10.3934/mbe.2023007
|
| [9] |
M. Abdar, M. A. Fahami, L. Rundo, P. Radeva, A. F. Frangi, U. R. Acharya, et al., Hercules: deep hierarchical attentive multilevel fusion model with uncertainty quantification for medical image classification, IEEE Trans. Ind. Inf., 19 (2023), 274–285. https://doi.org/10.1109/TII.2022.3168887 doi: 10.1109/TII.2022.3168887
|
| [10] |
Z. Yang, Y. Bao, Y. Liu, Q. Zhao, H. Zheng, Research on deep learning garbage classification system based on fusion of image classification and object detection classification, Math. Biosci. Eng., 20 (2022), 4741–4759. https://doi.org/10.3934/mbe.2023219 doi: 10.3934/mbe.2023219
|
| [11] |
H. Rastegar, D. Giveki, Designing a new deep convolutional neural network for skin lesion recognition, Multimedia Tools Appl., 82 (2023), 18907–18923. https://doi.org/10.1007/s11042-022-14181-4 doi: 10.1007/s11042-022-14181-4
|
| [12] |
Z. Wu, C. Liu, J. Wen, Y. Xu, J. Yang, X. Li, Selecting high-quality proposals for weakly supervised object detection with bottom-up aggregated attention and phase-aware loss, IEEE Trans. Image Process., 32 (2023), 682–693. https://doi.org/10.1109/TIP.2022.3231744 doi: 10.1109/TIP.2022.3231744
|
| [13] |
T. Diwan, G. Anirudh, J. V. Tembhurne, Object detection using yolo: challenges, architectural successors, datasets and applications, Multimedia Tools Appl., 82 (2023), 9243–9275. https://doi.org/10.1007/s11042-022-13644-y doi: 10.1007/s11042-022-13644-y
|
| [14] |
T. Shen, F. Huang, X. Zhang, CT medical image segmentation algorithm based on deep learning technology, Math. Biosci. Eng., 20 (2023), 10954–10976. https://doi.org/10.3934/mbe.2023485 doi: 10.3934/mbe.2023485
|
| [15] |
Y. Zhong, Z. Tang, H. Zhang, Y. Xie, X. Gao, A froth image segmentation method via generative adversarial networks with multi-scale self-attention mechanism, Multimedia Tools Appl., 2023. https://doi.org/10.1007/s11042-023-16397-4 doi: 10.1007/s11042-023-16397-4
|
| [16] |
S. Xue, H. Wang, X. Guo, M. Sun, K. Song, Y. Shao, et al., Cts-net: A segmentation network for glaucoma optical coherence tomography retinal layer images, Bioengineering, 10 (2023), 230. https://doi.org/10.3390/bioengineering10020230 doi: 10.3390/bioengineering10020230
|
| [17] |
H. Li, X. Tao, T. Liang, J. Jiang, J. Zhu, S. Wu, et al., Comprehensive ai-assisted tool for ankylosing spondylitis based on multicenter research outperforms human experts, Front. Public Health, 11 (2023), 1063633. https://doi.org/10.3389/fpubh.2023.1063633 doi: 10.3389/fpubh.2023.1063633
|
| [18] |
B. Cassidy, C. Kendrick, A. Brodzicki, J. Jaworek-Korjakowska, M. H. Yap, Analysis of the isic image datasets: usage, benchmarks and recommendations, Med. Image Anal., 75 (2022), 102305. https://doi.org/10.1016/j.media.2021.102305 doi: 10.1016/j.media.2021.102305
|
| [19] |
P. Tang, Q. Liang, X. Yan, S. Xiang, D. Zhang, Gp-cnn-dtel: Global-part cnn model with data-transformed ensemble learning for skin lesion classification, IEEE J. Biomed. Health. Inf., 24 (2020), 2870–2882. https://doi.org/10.1109/JBHI.2020.2977013 doi: 10.1109/JBHI.2020.2977013
|
| [20] |
X. He, Y. Wang, S. Zhao, C. Yao, Deep metric attention learning for skin lesion classification in dermoscopy images, Complex Intell. Syst., 8 (2022), 1487–1504. https://doi.org/10.1007/s40747-021-00587-4 doi: 10.1007/s40747-021-00587-4
|
| [21] |
F. Golnoori, F. Z. Boroujeni, A. Monadjemi, Metaheuristic algorithm based hyper-parameters optimization for skin lesion classification, Multimedia Tools Appl., 82 (2023), 25677–25709. https://doi.org/10.1007/s11042-023-14429-7 doi: 10.1007/s11042-023-14429-7
|
| [22] |
S. Ayas, Multiclass skin lesion classification in dermoscopic images using swin transformer model, Neural Comput. Appl., 35 (2023), 6713–6722. https://doi.org/10.1007/s00521-022-08053-z doi: 10.1007/s00521-022-08053-z
|
| [23] |
Z. Wei, Q. Li, H. Song, Dual attention based network for skin lesion classification with auxiliary learning, Biomed. Signal Process. Control, 74 (2022), 103549. https://doi.org/10.1016/j.bspc.2022.103549 doi: 10.1016/j.bspc.2022.103549
|
| [24] |
L. Wang, L. Zhang, X. Shu, Z. Yi, Intra-class consistency and inter-class discrimination feature learning for automatic skin lesion classification, Med. Image Anal., 85 (2023), 102746. https://doi.org/10.1016/j.media.2023.102746 doi: 10.1016/j.media.2023.102746
|
| [25] | M. Versaci, G. Angiulli, F. La Foresta, P. Crucitti, F. Lagana, D. Pellicano, et al., Innovative soft computing techniques for the evaluation of the mechanical stress state of steel plates, in Applied Intelligence and Informatics, (2022), 14–28. https://doi.org/10.1007/978-3-031-24801-6_2 |
| [26] |
D. Zhuang, K. Chen, J. M. Chang, Cs-af: A cost-sensitive multi-classifier active fusion framework for skin lesion classification, Neurocomputing, 491 (2022), 206–216. https://doi.org/10.1016/j.neucom.2022.03.042 doi: 10.1016/j.neucom.2022.03.042
|
| [27] |
Y. Wang, Y. Feng, L. Zhang, J. T. Zhou, Y. Liu, R. S. M. Goh, et al., Adversarial multimodal fusion with attention mechanism for skin lesion classification using clinical and dermoscopic images, Med. Image Anal., 81 (2022), 102535. https://doi.org/10.1016/j.media.2022.102535 doi: 10.1016/j.media.2022.102535
|
| [28] |
X. Deng, Q. Yin, P. Guo, Efficient structural pseudoinverse learning-based hierarchical representation learning for skin lesion classification, Complex Intell. Syst., 8 (2022), 1445–1457. https://doi.org/10.1007/s40747-021-00588-3 doi: 10.1007/s40747-021-00588-3
|
| [29] |
S. Zhou, S. Tian, L. Yu, W. Wu, D. Zhang, Z. Peng, et al., Fixmatch-ls: Semi-supervised skin lesion classification with label smoothing, Biomed. Signal Process. Control, 84 (2023), 104709. https://doi.org/10.1016/j.bspc.2023.104709 doi: 10.1016/j.bspc.2023.104709
|
| [30] |
Y. Wan, Y. Cheng, M. Shao, Mslanet: Multi-scale long attention network for skin lesion classification, Appl. Intell., 53 (2023), 12580–12598. https://doi.org/10.1007/s10489-022-03320-x doi: 10.1007/s10489-022-03320-x
|
| [31] |
K. A. Ogudo, R. Surendran, O. I. Khalaf, Optimal artificial intelligence based automated skin lesion detection and classification model, Comput. Syst. Sci. Eng., 44 (2023), 693–707. https://doi.org/10.32604/csse.2023.024154 doi: 10.32604/csse.2023.024154
|
| [32] |
W. X. Tsai, Y. C. Li, C. H. Lin, Skin lesion classification based on multi-model ensemble with generated levels-of-detail images, Biomed. Signal Process. Control, 85 (2023), 105068. https://doi.org/10.1016/j.bspc.2023.105068 doi: 10.1016/j.bspc.2023.105068
|
| [33] |
F. Miao, L. Yao, X. Zhao, Adaptive margin aware complement-cross entropy loss for improving class imbalance in multi-view sleep staging based on eeg signals, IEEE Trans. Neural Syst. Rehabil. Eng., 30 (2022), 2927–2938. https://doi.org/10.1109/TNSRE.2022.3213848 doi: 10.1109/TNSRE.2022.3213848
|
| [34] |
D. Zabihzadeh, A. Tuama, A. Karami-Mollaee, S. J. Mousavirad, Low-rank robust online distance/similarity learning based on the rescaled hinge loss, Appl. Intell., 53 (2023), 634–657. https://doi.org/10.1007/s10489-022-03419-1 doi: 10.1007/s10489-022-03419-1
|
| [35] | T. Y. Lin, P. Goyal, R. Girshick, K. He, P. Dollár, Focal loss for dense object detection, in Proceedings of the IEEE International Conference on Computer Vision (ICCV), (2017), 2980–2988. |
| [36] |
G. Yue, P. Wei, T. Zhou, Q. Jiang, W. Yan, T. Wang, Toward multicenter skin lesion classification using deep neural network with adaptively weighted balance loss, IEEE Trans. Med. Imaging, 42 (2022), 119–131. https://doi.org/10.1109/TMI.2022.3204646 doi: 10.1109/TMI.2022.3204646
|
| [37] |
J. Du, K. Guan, P. Liu, Y. Li, T. Wang, Boundary-sensitive loss function with location constraint for hard region segmentation, IEEE J. Biomed. Health. Inf., 27 (2022), 992–1003. https://doi.org/10.1109/JBHI.2022.3222390 doi: 10.1109/JBHI.2022.3222390
|
| [38] |
K. Chaitanya, E. Erdil, N. Karani, E. Konukoglu, Local contrastive loss with pseudo-label based self-training for semi-supervised medical image segmentation, Med. Image Anal., 87 (2023), 102792. https://doi.org/10.1016/j.media.2023.102792 doi: 10.1016/j.media.2023.102792
|
| [39] | K. He, X. Zhang, S. Ren, J. Sun, Deep residual learning for image recognition, in Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition (CVPR), (2016), 770–778. |
| [40] | M. Tan, Q. Le, Efficientnetv2: Smaller models and faster training, in Proceedings of the 38th International Conference on Machine Learning, 139 (2021), 10096–10106. |
| [41] | Z. Liu, H. Mao, C. Y. Wu, C. Feichtenhofer, T. Darrell, S. Xie, A convnet for the 2020s, in Proceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition (CVPR), (2022), 11976–11986. |
| [42] |
X. Wu, Z. Tao, B. Jiang, T. Wu, X. Wang, H. Chen, Domain knowledge-enhanced variable selection for biomedical data analysis, Inf. Sci., 606 (2022), 469–488. https://doi.org/10.1016/j.ins.2022.05.076 doi: 10.1016/j.ins.2022.05.076
|
| [43] | A. Marghoob, R. Braun, An Atlas of Dermoscopy, CRC Press, 2012. https://doi.org/10.3109/9781841847627 |
| [44] | F. Schroff, D. Kalenichenko, J. Philbin, Facenet: A unified embedding for face recognition and clustering, in 2015 IEEE Conference on Computer Vision and Pattern Recognition (CVPR), (2015), 815–823. https://doi.org/10.1109/CVPR.2015.7298682 |
| [45] |
P. Tschandl, C. Rosendahl, H. Kittler, The ham10000 dataset, a large collection of multi-source dermatoscopic images of common pigmented skin lesions, Sci. Data, 5 (2018), 180161. https://doi.org/10.1038/sdata.2018.161 doi: 10.1038/sdata.2018.161
|
| [46] | M. Combalia, N. C. F. Codella, V. Rotemberg, B. Helba, V. Vilaplana, O. Reiter, et al., Bcn20000: Dermoscopic lesions in the wild, preprint, arXiv: 1908.02288. |
| [47] | J. Deng, W. Dong, R. Socher, L. J. Li, K. Li, F. F. Li, Imagenet: A large-scale hierarchical image database, in 2009 IEEE Conference on Computer Vision and Pattern Recognition, (2009), 248–255. https://doi.org/10.1109/CVPR.2009.5206848 |
| [48] | Y. Cui, M. Jia, T. Y. Lin, Y. Song, S. Belongie, Class-balanced loss based on effective number of samples, in Proceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition (CVPR), (2019), 9268–9277. |








Figures(9) / Tables(4)

Li Zhang, Xiangling Xiao, Ju Wen, Huihui Li. MDKLoss: Medicine domain knowledge loss for skin lesion recognition[J]. Mathematical Biosciences and Engineering, 2024, 21(2): 2671-2690. doi: 10.3934/mbe.2024118







 DownLoad:
DownLoad: