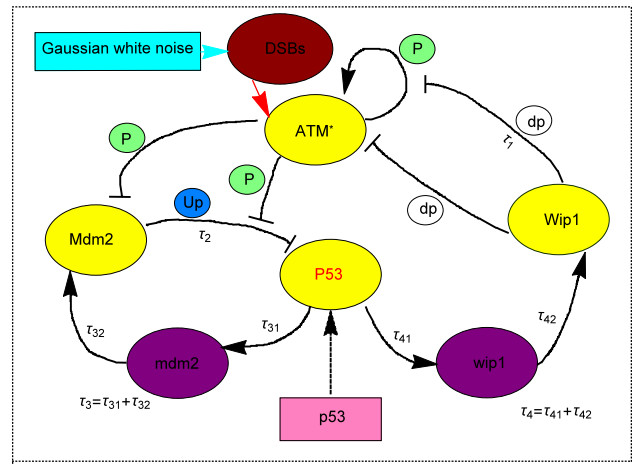
The tumor suppressor protein P53 can regulate the cell cycle, thereby preventing cell abnormalities. In this paper, we study the dynamic characteristics of the P53 network under the influence of time delay and noise, including stability and bifurcation. In order to study the influence of several factors on the concentration of P53, bifurcation analysis on several important parameters is conducted; the results show that the important parameters could induce P53 oscillations within an appropriate range. Then we study the stability of the system and the existing conditions of Hopf bifurcation by using Hopf bifurcation theory with time delays as the bifurcation parameter. It is found that time delay plays a key role in inducing Hopf bifurcation and regulating the period and amplitude of system oscillation. Meanwhile, the combination of time delays can not only promote the oscillation of the system but it also provides good robustness. Changing the parameter values appropriately can change the bifurcation critical point and even the stable state of the system. In addition, due to the low copy number of the molecules and the environmental fluctuations, the influence of noise on the system is also considered. Through numerical simulation, it is found that noise not only promotes system oscillation but it also induces system state switching. The above results may help us to further understand the regulation mechanism of the P53-Mdm2-Wip1 network in the cell cycle.
Citation: LanJiang Luo, Haihong Liu, Fang Yan. Dynamic behavior of P53-Mdm2-Wip1 gene regulatory network under the influence of time delay and noise[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 2321-2347. doi: 10.3934/mbe.2023109
The tumor suppressor protein P53 can regulate the cell cycle, thereby preventing cell abnormalities. In this paper, we study the dynamic characteristics of the P53 network under the influence of time delay and noise, including stability and bifurcation. In order to study the influence of several factors on the concentration of P53, bifurcation analysis on several important parameters is conducted; the results show that the important parameters could induce P53 oscillations within an appropriate range. Then we study the stability of the system and the existing conditions of Hopf bifurcation by using Hopf bifurcation theory with time delays as the bifurcation parameter. It is found that time delay plays a key role in inducing Hopf bifurcation and regulating the period and amplitude of system oscillation. Meanwhile, the combination of time delays can not only promote the oscillation of the system but it also provides good robustness. Changing the parameter values appropriately can change the bifurcation critical point and even the stable state of the system. In addition, due to the low copy number of the molecules and the environmental fluctuations, the influence of noise on the system is also considered. Through numerical simulation, it is found that noise not only promotes system oscillation but it also induces system state switching. The above results may help us to further understand the regulation mechanism of the P53-Mdm2-Wip1 network in the cell cycle.

| [1] |
E. Batchelor, A. Loewer, Recent progress and open challenges in modeling p53 dynamics in single cells, Curr. Opin. Syst. Biol., 3 (2017), 54–59. https://doi.org/10.1016/j.coisb.2017.04.007 doi: 10.1016/j.coisb.2017.04.007

|
| [2] |
K. H. Vousden, D. P. Lane, P53 in health and disease, Nat. Rev. Mol. Cell Biol., 8 (2007), 275–283. https://doi.org/10.1038/nrm2147 doi: 10.1038/nrm2147
|
| [3] |
P. A. J. Muller, K. H. Vousden, P53 mutations in cancer, Nat. Cell Biol., 15 (2013), 2–8. https://doi.org/10.1038/ncb2641 doi: 10.1038/ncb2641
|
| [4] |
X. Wu, J. H. Bayle, D. Olson, A. J. Levine, The p53-Mdm2 autoregulatory feedback loop, Genes Dev., 7 (1993), 1126–1132. https://doi.org/10.1101/gad.7.7.1126 doi: 10.1101/gad.7.7.1126
|
| [5] |
T. Sun, J. Cui, Dynamics of P53 in response to DNA damage: Mathematical modeling and perspective, Prog. Biophys. Mol. Biol., 119 (2015), 175–182. https://doi.org/10.1016/j.pbiomolbio.2015.08.017 doi: 10.1016/j.pbiomolbio.2015.08.017
|
| [6] |
A. Loewer, E. Batchelor, G. Gaglia, G. Lahav, Basal dynamics of p53 reveal transcriptionally attenuated pulses in cycling cells, Cell, 142 (2010), 89–100. https://doi.org/10.1016/j.cell.2010.05.031 doi: 10.1016/j.cell.2010.05.031
|
| [7] |
G. $M\ddot{o}nke$, E. Cristiano, A. Finzel, Excitability in the p53 network mediates robust signaling with tunable activation thresholds in single cells, Sci. Rep., 7 (2017), 46571. https://doi.org/10.1038/srep46571 doi: 10.1038/srep46571
|
| [8] |
E. R. Kastenhuber, S. W. Lowe, Putting p53 in context, Cell, 170 (2017), 1062–1078. https://doi.org/10.1016/j.cell.2017.08.028 doi: 10.1016/j.cell.2017.08.028
|
| [9] |
J. Stewart-Ornstein, G. Lahav, P53 dynamics in response to DNA damage vary across cell lines and are shaped by efficiency of DNA repair and activity of the kinase ATM, Sci. Signaling, 10 (2017), eaah6671. https://doi.org/10.1126/scisignal.aah6671 doi: 10.1126/scisignal.aah6671
|
| [10] |
A, J. Levine, P53, the cellular gatekeeper for growth and division, Cell, 88 (1997), 323–331. https://doi.org/10.1016/s0092-8674(00)81871-1 doi: 10.1016/s0092-8674(00)81871-1
|
| [11] |
J. E. Purvis, K. W. Karhohs, C. Mock, P53 dynamics control cell fate, Science, 336 (2012), 1440–1444. https://doi.org/10.1126/science.1218351 doi: 10.1126/science.1218351
|
| [12] |
C. Gao, J. Ji, F. Yan, H. Liu, Oscillation induced by Hopf bifurcation in the p53$-$Mdm2 feedback module, IET Syst. Biol., 13 (2019), 251–259. https://doi.org/10.1049/iet-syb.2018.5092 doi: 10.1049/iet-syb.2018.5092
|
| [13] |
J. D. Olinger, K. W. Kinzler, P. S Meltzer, D. L. George, Amplification of a gene encoding a p53$-$associated protein in human sarcomas, Nature, 358 (1992), 80–83. https://doi.org/10.1038/358080a0 doi: 10.1038/358080a0
|
| [14] |
M. H. Kubbutat, S. N. Jones, K, H. Vousden, Regulation of p53 stability by Mdm2, Nature, 387 (1997), 299–303. https://doi.org/10.1038/387299a0 doi: 10.1038/387299a0
|
| [15] |
J. H. Park, S. W. Yang, J. M. Park, Positive feedback regulation of p53 transactivity by DNA damage-induced ISG15 modification, Nat. Commun., 7 (2016), 12513. https://doi.org/10.1038/ncomms12513 doi: 10.1038/ncomms12513
|
| [16] |
K. H. Vousden, D. P. Lane, P53 in health and disease, Nat. Rev. Mol. Cell Biol., 8 (2007), 275–283. https://doi.org/10.1038/nrm2147 doi: 10.1038/nrm2147
|
| [17] |
N. D. Lakin, S. P. Jackson, Regulation of p53 in response to DNA damage, Oncogene, 18 (1999), 7644–7655. https://doi.org/10.1038/sj.onc.1203015 doi: 10.1038/sj.onc.1203015
|
| [18] |
G. Wang, Z. Yang, Stability and Hopf bifurcation analysis in a delayed Myc/E2F/miR-17-92 network involving interlinked positive and negative feedback loops, Discrete Dyn. Nat. Soc., 2018 (2018), 7014789. https://doi.org/10.1155/2018/7014789 doi: 10.1155/2018/7014789
|
| [19] |
B. D. Aguda, Y. Kim, M. G. P. Hunter, A. Friedman, C. B. Marsh, MicroRNA regulation of a cancer network: Consequences of the feedback loops involving miR-17-92, E2F, and Myc, Proc. Acad. Sci., 105 (2008), 19678–19683. https://doi.org/10.1073/pnas.0811166106 doi: 10.1073/pnas.0811166106
|
| [20] |
Y. Zhang, H. Liu, F. Yan, J. Zhou, Oscillatory dynamics of p38 activity with transcriptional and translational time delays, Sci. Rep., 7 (2017), e11495l. https://doi.org/10.1038/s41598-017-11149-5 doi: 10.1038/s41598-017-11149-5
|
| [21] |
M. Chen, J. Ji, H. Liu, F. Yan, Periodic oscillations in the quorum-sensing system with time delay, Int. J. Bifurcation Chaos, 30 (2020), 2050127. https://doi.org/10.1142/S0218127420501278 doi: 10.1142/S0218127420501278
|
| [22] |
N. Monk, Oscillatory expression of Hes1, p53, and NF-kappaB driven by transcriptional time delays, Curr. Biol., 13 (2003), 1409–1413. https://doi.org/10.1016/s0960-9822(03)00494-9 doi: 10.1016/s0960-9822(03)00494-9
|
| [23] |
P. He, K. J. Billy, H. Ma, Y. Jia, L. Yang, The dynamical roles of miR-17-92 on the E2F-related network during the G1/Stransition, Nonlinear Dyn., 95 (2019), 259–271. https://doi.org/10.1007/s11071-018-4562-x doi: 10.1007/s11071-018-4562-x
|
| [24] | Y. Ding, J. Shen, J. Lu, J. Kurths, Stochastic resonance in genetic regulatory networks under L'evy noise, Europhys. Lett., 127 (2019), 50003. |
| [25] |
E. M. Ozbudak, M. Thattal, N. L. Han, Multistability in the lactose utilization network of Escherichia coli, Nature, 427 (2004), 737–740. https://doi.org/10.1038/nature02298 doi: 10.1038/nature02298
|
| [26] |
C. J. Bakkenist, M. B. Kastan, DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation, Nature, 421 (2003), 499–506. https://doi.org/10.1038/nature01368 doi: 10.1038/nature01368
|
| [27] |
M. F. Lavin, Ataxia-telangiectasia: From a rare disorder to a paradigm for cell signalling and cancer, Nat. Rev. Mol. Cell Biol., 9 (2008), 759–769. https://doi.org/10.1038/nrm2514 doi: 10.1038/nrm2514
|
| [28] |
A. Ma, X. Dai, Exploring the influence of parameters on the p53 response when single-stranded breaks and double-stranded breaks coexist, Interdiscip. Sci. Comput. Life Sci., 11 (2019), 679–690. https://doi.org/10.1007/s12539-019-00332-z doi: 10.1007/s12539-019-00332-z
|
| [29] |
Y. T. Barak, T. Juven, R. Haffner, M. Oren, MDM2 expression is induced by wild type p53 activity, EMBO J., 12 (1993), 461–468. https://doi.org/10.1002/j.1460-2075.1993.tb05678.x doi: 10.1002/j.1460-2075.1993.tb05678.x
|
| [30] |
M. L. Heltberg, S. H. Chen, A. Jimenez, G. Lahav, Inferring leading interactions in the p53/Mdm2/Mdmx circuit through live-cell imaging and modeling, Cell Syst., 9 (2019), 548–558. https://doi.org/10.1016/j.cels.2019.10.010 doi: 10.1016/j.cels.2019.10.010
|
| [31] |
R. Yang, B. Huang, Y. Zhu, Y. Li, F. Liu, J. Shi, Cell type-dependent bimodal p53 activation engenders a dynamic mechanism of chemoresistance, Sci. Adv., 4 (2018), eaat5077. https://doi.org/10.1126/sciadv.aat5077 doi: 10.1126/sciadv.aat5077
|
| [32] |
K. Karanam, K. Ran, A. Loewer, Quantitative live cell imaging reveals a gradual shift between DNA repair mechanisms and a maximal use of HR in mid S phase, Mol. Cell, 47 (2012), 320–329. https://doi.org/10.1016/j.molcel.2012.05.052 doi: 10.1016/j.molcel.2012.05.052
|
| [33] | D. T. Gillespie, Markov Processes: An Introduction for Physical Scientists, Centre Dedocumentation Universitaire Cdu, 34 (1992), 87. https://doi.org/10.1016/0378-4754(92)90114-V |
| [34] |
W. W. Yu, J. D. Cao, Stability and Hopf bifurcation analysis on a four-neuron BAM neural network with time delays, Phys. Lett. A, 351 (2006), 64–78. https://doi.org/10.1016/j.physleta.2005.10.056 doi: 10.1016/j.physleta.2005.10.056
|
| [35] |
D. T. Gillespie, The chemical Langevin equation, J. Chem. Phys., 113 (2000), 297–306. https://doi.org/10.1063/1.481811 doi: 10.1063/1.481811
|
| [36] | H. Hu, Z. Wang, Dynamics of Controlled Mechanical Systems with Delayed Feedback, Springer Heidelberg, 2002. https://doi.org/10.1007/978-3-662-05030-9 |
| [37] |
S. Ruan, J. Wei, On the zeros of transcendental functions with applications to stability of delayed differential equations with two delays, Dyn. Contin. Discrete Impulsive Syst., 10 (2003), 863–874. https://doi.org/10.1093/imammb/18.1.41 doi: 10.1093/imammb/18.1.41
|
| [38] |
I. Mendez-Balbuena, P. Arrieta, N. Huidobro, Augmenting EEG-global-coherence with auditory and visual noise, Medicine, 97 (2018), 863–874. https://doi.org/10.1097/MD.0000000000012008 doi: 10.1097/MD.0000000000012008
|
| [39] |
P. Lai, T. Y. Chi, G. G. Chen, Different levels of p53 induced either apoptosis or cell cycle arrest in a doxycycline-regulated hepatocellular carcinoma cell line in vitro, Apoptosis, 12 (2007), 387–393. https://doi.org/10.1007/s10495-006-0571-1 doi: 10.1007/s10495-006-0571-1
|
| [40] |
K. H. Chong, X. Zhang, Z. Jie, Dynamical analysis of cellular ageing by modeling of gene regulatory network based attractor landscape, Plos One, 13 (2018), e0197838. https://doi.org/10.1371/journal.pone.0197838 doi: 10.1371/journal.pone.0197838
|
| [41] |
Y. Cao, A. Lopatkin, L. You, Elements of biological oscillations in time and space, Nat. Struct. Mol. Biol., 2016 (2016). https://doi.org/10.1038/nsmb.3320 doi: 10.1038/nsmb.3320
|
| [42] |
M. $Kocha\acute{n}czyk$, P. Kocieniewski, E. Kozłowska, Relaxation oscillations and hierarchy of feedbacks in MAPK signaling, Sci. Rep., 7 (2017), 1–15. https://doi.org/10.1038/srep38244 doi: 10.1038/srep38244
|
| [43] |
B. Hat, K. Puszynski, T. Lipniacki, Exploring mechanisms of oscillations in p53 and nuclear factor-B systems, IET Syst. Biol., 3 (2009), 342–355. https://doi.org/10.1049/iet-syb.2008.0156 doi: 10.1049/iet-syb.2008.0156
|
| [44] |
C. Zeng, T. Yang, Q. Han, C. Zhang, D. Tian, H. Wang, Noises-induced toggle switch and stability in a gene regulation network, Int. J. Modern Phys. B, 28 (2014), e1450223. https://doi.org/10.1142/S0217979214502233 doi: 10.1142/S0217979214502233
|
| [45] |
H. Zhang, Y. Chen, Noise propagation in gene regulation networks involving interlinked positive and negative feedback loops, Plos One, 7 (2012), e51840. https://doi.org/10.1371/journal.pone.0051840 doi: 10.1371/journal.pone.0051840
|
| [46] |
P. Zhou, S. Cai, Z. Liu, R. Wang, Mechanisms generating bistability and oscillations in microRNA-mediated motifs, Phys. Rev. E, 85 (2012), 041916. https://doi.org/10.1103/PhysRevE.85.041916 doi: 10.1103/PhysRevE.85.041916
|
| [47] |
V. Chickarmane, A. Ray, H. M. Sauro, A. Nadim, A model for p53 dynamics triggered by DNA damage, SIAM J. Appl. Dyn. Syst., 6 (2007), 61–78. https://doi.org/10.1137/060653925 doi: 10.1137/060653925
|
| [48] |
D. G. Wang, S. Wang, B. Huang, F. Liu, Roles of cellular heterogeneity, intrinsic and extrinsic noise in variability of p53 oscillation, Sci. Rep., 9 (2019), 1–11. https://doi.org/10.1038/s41598-019-41904-9 doi: 10.1038/s41598-019-41904-9
|
| [49] |
Y. C. Hung, C. K. Hu, Constructive role of noise in p53 regulatory network, Comput. Phys. Commun., 182 (2011), 249–250. https://doi.org/10.1016/j.cpc.2010.07.002 doi: 10.1016/j.cpc.2010.07.002
|
| [50] |
S. Ditlevsen, P. Greenwood, The MorrisCLecar neuron model embeds a leaky integrate-and-fire model, J. Math. Biol., 67 (2011), 239–259. https://doi.org/10.1007/s00285-012-0552-7 doi: 10.1007/s00285-012-0552-7
|
| [51] |
N. Yang, T. Sun, P. Shen, Deciphering p53 dynamics and cell fate in DNA damage response using mathematical modeling, Genome Instab. Dis., 1 (2020), 265–277. https://doi.org/10.1007/s42764-020-00019-6 doi: 10.1007/s42764-020-00019-6
|
| [52] |
L. Ma, J. Wagner, J. J. Rice, W. Hu, A. J. Levine, G. A. Stolovitzky, A plausible model for the digital response of p53 to DNA damage, Proc. Natl. Acad. Sci. USA, 102 (2005), 14266–14271. https://doi.org/10.1073/pnas.0501352102 doi: 10.1073/pnas.0501352102
|
| [53] |
E. Batchelor, C. Mock, I. Bhan, A. Loewer, G. Lahav, Recurrent initiation: A mechanism for triggering p53 pulses in response to DNA damage, Mol. Cell, 30 (2008), 277–289. https://doi.org/10.1016/j.molcel.2008.03.016 doi: 10.1016/j.molcel.2008.03.016
|
| [54] |
X. Zhang, F. Liu, W. Wang, Two-phase dynamics of p53 in the dna damage response, Proc. Natl. Acad. Sci. USA, 108 (2011), 8990-8995. https://doi.org/10.1073/pnas.1100600108 doi: 10.1073/pnas.1100600108
|
| [55] |
K. $Puszy\acute{n}ski$, B. Hat, T. Lipniacki, Oscillations and bistability in the stochastic model of p53 regulation, J. Theor. Biol., 254 (2008), 452–465. https://doi.org/10.1016/j.jtbi.2008.05.039 doi: 10.1016/j.jtbi.2008.05.039
|
| [56] |
T. Sun, R. Yuan, W. Xu, F. Zhu, P. Shen, Exploring a minimal two-component p53 model, Phys. Biol., 7 (2010), 36008–36008. https://doi.org/10.1088/1478-3975/7/3/036008 doi: 10.1088/1478-3975/7/3/036008
|















Figures(16) / Tables(1)

LanJiang Luo, Haihong Liu, Fang Yan. Dynamic behavior of P53-Mdm2-Wip1 gene regulatory network under the influence of time delay and noise[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 2321-2347. doi: 10.3934/mbe.2023109







 DownLoad:
DownLoad: