Citation: Sergei Trofimchuk, Vitaly Volpert. Traveling waves in delayed reaction-diffusion equations in biology[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6487-6514. doi: 10.3934/mbe.2020339

| [1] | A. Kolmogorov, I. Petrovskii, N. Piskunov, Study of a diffusion equation that is related to the growth of a quality of matter and its application to a biological problem, Byul. Mosk. Gos. Univ. Ser. A Mat. Mekh., 1 (1937) 1-26. |
| [2] |
A. Friedman, Cancer as multifaceted disease, Math. Model. Nat. Phenom., 7 (2012), 3-28. doi: 10.1051/mmnp/20127102

|
| [3] | A. V. Panfilov, H. Dierckx, V. Volpert, Reaction-diffusion waves in cardiovascular diseases, Phys. D Amsterdam, Neth. 399 (2019), 1-34. |
| [4] | J. Bebernes, D. Eberly, Mathematical Problems from Combustion Theory, Springer-Verlag, New York, (1989). |
| [5] | R. A. Fisher, The wave of advance of advantageous genes, Ann. Hum. Genet., 7 (1937), 353-369. |
| [6] | Y. B. Zeldovich, D. A. Frank-Kamenetskii, A theory of thermal propagation of flame, Acta Physicochim. USSR, 9 (1938), 341-350. |
| [7] | N. N. Semenov, To the theory of combustion processes, Fiz. Khim., 4 (1939), 4-7. |
| [8] | V. A. Mikhelson, On normal combustion velocity of explosive gaseous mixtures, Imp. Moscow Univ. Sci. Bull. Phys. Math. Ser., 10 (1893), 1-92. |
| [9] | R. Luther, Propagation of chemical reactions in space, J. Chem. Educ., 64 (9), 740. |
| [10] |
J. Xin, Front propagation in heterogeneous media, SIAM Rev., 42 (2000), 161-230. doi: 10.1137/S0036144599364296
|
| [11] | A. I. Volpert, V. A. Volpert, V. A. Volpert, Traveling Wave Solutions of Parabolic Systems, in American Mathematical Society, 1994. |
| [12] | A. I. Volpert, V. A. Volpert, Application of the theory of the rotation of vector fields to the investigation of wave solutions of parabolic equations, Proc. Moscow Math. Soc., 52 (1989), 58-109. |
| [13] |
P. C. Fife, J. B. McLeod, The approach of solutions of nonlinear diffusion equations to travelling front solutions, Arch. Ration. Mech. Anal., 65 (1977), 335-361. doi: 10.1007/BF00250432
|
| [14] |
P. C. Fife, J. B. McLeod, A phase plane discussion of convergence to travelling fronts for nonlinear diffusion, Arch. Ration. Mech. Anal., 75 (1981), 281-314. doi: 10.1007/BF00256381
|
| [15] |
V. A. Volpert, Asymptotic behavior of solutions of a nonlinear diffusion equation with a source of general type, Sib. Math. J., 30 (1989), 25-36. doi: 10.1007/BF01054212
|
| [16] |
V. A. Volpert, Convergence to a wave of solutions of a nonlinear diffusion equation with a source of general type, Sib. Math. J., 30 (1989), 203-210. doi: 10.1007/BF00971374
|
| [17] | M. Artola, Equations paraboliques à retardement, C. R. Acad. Sci. Paris, 264 (1967), 668-671. |
| [18] | M. Artola, Sur les perturbations des équations d'évolution: Application à des problèmes de retard, Annales scientifiques de l'école Normale Supérieure, Serie 4, 1969. Available from: http://www.numdam.org/article/ASENS_1969_4_2_2_137_0. |
| [19] |
C. C. Travis, G. F. Webb, Existence and stability for partial functional differential equations, Trans. Amer. Math. Soc., 200 (1974), 395-418. doi: 10.1090/S0002-9947-1974-0382808-3
|
| [20] | R. H. Martin, H. L. Smith, Abstract functional differential equations and reaction-diffusion systems, Trans. Amer. Math. Soc., 321 (1990), 1-44. |
| [21] | J. Wu, Theory and Applications of Partial Functional Differential Equations, Springer-Verlag, 1996. |
| [22] | K. Schumacher, Travelling-front solutions for integro-differential equations I, J. Reine Angew. Math., 316 (1980), 54-70. |
| [23] | K. Schaaf, Asymptotic behavior and travelling wave solutions for parabolic functional differential equations, Trans. Amer. Math. Soc., 302 (1987), 587-615. |
| [24] |
R. Benguria, A. Solar, An iterative estimation for disturbances of semi-wavefronts to the delayed Fisher-KPP equation, Proc. Amer. Math. Soc., 147 (2019), 2495-2501. doi: 10.1090/proc/14381
|
| [25] |
J. Billingham, Slow travelling wave solutions of the nonlocal Fisher-KPP equation, Nonlinearity, 33 (2020), 2106-2142. doi: 10.1088/1361-6544/ab6f4f
|
| [26] |
A. Ducrot, G. Nadin, Asymptotic behaviour of traveling waves for the delayed Fisher-KPP equation, J. Differ. Equations, 256 (2014), 3115-3140. doi: 10.1016/j.jde.2014.01.033
|
| [27] |
K. Hasík, S. Trofimchuk, An extension of Wright's 3/2-theorem for the KPP-Fisher delayed equation, Proc. Amer. Math. Soc., 143 (2015), 3019-3027. doi: 10.1090/S0002-9939-2015-12496-3
|
| [28] |
E. Hernández, S. Trofimchuk, Nonstandard quasi-monotonicity: an application to the wave existence in a neutral KPP-Fisher equation, J. Dynam. Differ. Equations, 32 (2020), 921-939. doi: 10.1007/s10884-019-09748-z
|
| [29] |
Y. Liu, P. Weng, Asymptotic pattern for a partial neutral functional differential equation, J. Differ. Equations, 258 (2015), 3688-3741. doi: 10.1016/j.jde.2015.01.016
|
| [30] | S. Ma, X. Zou, Existence, uniqueness and stability of travelling waves in a discrete reactiondiffusion monostable equation with delay, J. Differ. Equations, 217 (2005), 54-87. |
| [31] |
S. A. Gourley, J. W. H. So, J.H. Wu, Non-locality of reaction-diffusion equations induced by delay: Biological modeling and nonlinear dynamics, J. Math. Sci., 124 (2004), 5119-5153. doi: 10.1023/B:JOTH.0000047249.39572.6d
|
| [32] | J. D. Murray, Mathematical Biology, Vol. 1, 3rd edition, Springer-Verlag, (2002). |
| [33] |
M. Aguerrea, C. Gomez, S. Trofimchuk, On uniqueness of semi-wavefronts Diekmann-Kaper theory of a nonlinear convolution equation re-visited, Math. Ann., 354 (2012), 73-109. doi: 10.1007/s00208-011-0722-8
|
| [34] | J. Mallet-Paret, The Fredholm alternative for functional differential equations of mixed type, J. Dyn. Differ. Equations, 11 (1999), 1-48. |
| [35] |
E. Trofimchuk, M. Pinto, S. Trofimchuk, Pushed traveling fronts in monostable equations with monotone delayed reaction, Discrete Contin. Dyn. Syst. A, 33 (2013), 2169-2187. doi: 10.3934/dcds.2013.33.2169
|
| [36] | T. Xu, S. Ji, R. Huang, M. Mei, J. Yin, Theoretical and numerical studies on global stability of traveling waves with oscillations for time-delayed nonlocal dispersion equations, Int. J. Numer. Anal. Mod., 17 (2020), 68-86. |
| [37] | T. Yi, X. Zou, Asymptotic behavior, spreading speeds, and traveling waves of nonmonotone dynamical systems, SIAM J. Math. Anal., 47 (2015), 3005-3034. |
| [38] |
N. F. Britton, Aggregation and the competitive exclusion principle, J. Theor. Biol., 136 (1989), 57-66. doi: 10.1016/S0022-5193(89)80189-4
|
| [39] |
N. F. Britton, Spatial structures and periodic travelling waves in an integro- differential reactiondiffusion population model, SIAM J. Appl. Math., 50 (1990), 1663-1688. doi: 10.1137/0150099
|
| [40] |
H. L. Smith, A structured population model and a related functional-differential equation: Global attractors and uniform persistence, J. Dyn. Differ. Equations, 6 (1994), 71-99. doi: 10.1007/BF02219189
|
| [41] |
H. L. Smith, H. Thieme, Strongly order preserving semiflows generated by functional differential equations, J. Differ. Equations, 93 (1991), 332-363. doi: 10.1016/0022-0396(91)90016-3
|
| [42] |
J. W. H. So, J. H. Wu, X. F. Zou, A reaction-diffusion model for a single species with age structure I: Traveling wavefronts on unbounded domains, Proc. R. Soc. London, Ser. A, 457 (2001), 1841-1853. doi: 10.1098/rspa.2001.0789
|
| [43] |
M. Bani-Yaghoub, G. M. Yao, M. Fujiwara, D.E. Amundsen, Understanding the interplay between density dependent birth function and maturation time delay using a reaction-diffusion population model, Ecol. Complexity, 21 (2015), 14-26. doi: 10.1016/j.ecocom.2014.10.007
|
| [44] |
V. Volpert, S. Petrovskii, Reaction-diffusion waves in biology, Phys. Life Rev., 6 (2009), 267-310. doi: 10.1016/j.plrev.2009.10.002
|
| [45] | X. Chen, Existence, uniqueness, and asymptotic stability of traveling waves in nonlocal evolution equations, Adv. Differ. Equations, 2 (1997), 125-160. |
| [46] |
T. Ogiwara, H. Matano, Monotonicity and convergence in order-preserving systems, Discrete Contin. Dyn. Syst. A, 5 (1999), 1-34. doi: 10.3934/dcds.1999.5.1
|
| [47] |
J. Fang, X. Q. Zhao, Bistable traveling waves for monotone semiflows with applications, J. Eur. Math. Soc., 17 (2015), 2243-2288. doi: 10.4171/JEMS/556
|
| [48] |
J. Fang, X.-Q. Zhao, Traveling waves for monotone semiflows with weak compactness, SIAM J. Math. Anal., 46 (2014), 3678-3704. doi: 10.1137/140953939
|
| [49] |
T. Faria, W. Huang, J. Wu, Traveling waves for delayed reaction-diffusion equations with nonlocal response, Proc. R. Soc. Ser. A, 462 (2006), 229-261. doi: 10.1098/rspa.2005.1554
|
| [50] |
X. Liang, X.-Q. Zhao, Spreading speeds and traveling waves for abstract monostable evolution systems, J. Funct. Anal., 259 (2010), 857-903. doi: 10.1016/j.jfa.2010.04.018
|
| [51] |
H. Yagisita, Existence and nonexistence of traveling waves for a nonlocal monostable equation, Publ. Res. Inst. Math. Sci., 45 (2009), 925-953. doi: 10.2977/prims/1260476648
|
| [52] |
H. Yagisita, Existence of traveling waves solutions for a nonlocal bistable equation:an abstract approach, Publ. RIMS Kyoto Univ., 45 (2009), 955-979. doi: 10.2977/prims/1260476649
|
| [53] | W. van Saarloos, Front propagation into unstable states, Phys. Rep., 386 (2003), 29-222. |
| [54] | B. Sandstede, Stability of travelling waves, in Handbook of dynamical systems Ⅱ (eds. B. Fiedler), Elsevier, (2002), 983-1055. |
| [55] |
I. L. Chern, M. Mei, X. F. Yang, Q. F. Zhang, Stability of non-monotone critical traveling waves for reaction-diffusion equations with time-delay, J. Differ. Equations, 259 (2015), 1503-1541. doi: 10.1016/j.jde.2015.03.003
|
| [56] |
C. K. Lin, C. T. Lin, Y. Lin, M. Mei, Exponential stability of nonmonotone traveling waves for Nicholson's blowflies equation, SIAM J. Math. Anal., 46 (2014), 1053-1084. doi: 10.1137/120904391
|
| [57] |
M. Mei, C. K. Lin, C. T. Lin, J. W. H. So, Traveling wavefronts for time-delayed reactiondiffusion equation: (I) Local nonlinearity, J. Differ. Equations, 247 (2009), 495-510. doi: 10.1016/j.jde.2008.12.026
|
| [58] |
M. Mei, C. K. Lin, C. T. Lin, J. W.-H. So, Traveling wavefronts for time-delayed reactiondiffusion equation: (Ⅱ) Nonlocal nonlinearity, J. Differ. Equations, 247 (2009), 511-529. doi: 10.1016/j.jde.2008.12.020
|
| [59] |
M. Mei, J. W. H. So, M. Y. Li, S. S. P. Shen, Asymptotic stability of traveling waves for the Nicholson's blowflies equation with diffusion, Proc. R. Soc. Edinburgh, Sect. A, 134 (2004), 579-594. doi: 10.1017/S0308210500003358
|
| [60] | M. Mei, Y. Wang, Remark on stability of traveling waves for nonlocal Fisher-KPP equations, Int. J. Numer. Anal. Model. Ser. B, 2 (2011), 379-401. |
| [61] |
G. Lv, M. Wang, Nonlinear stability of travelling wave fronts for delayed reaction diffusion equations, Nonlinearity, 23 (2010), 845-873. doi: 10.1088/0951-7715/23/4/005
|
| [62] |
S. L. Wu, W. T. Li, S. Y. Liu, Exponential stability of traveling fronts in monostable reactionadvection-diffusion equations with non-local delay, Discrete Contin. Dyn. Syst. B, 17 (2012), 347-366. doi: 10.3934/dcdsb.2012.17.347
|
| [63] |
A. Solar, S. Trofimchuk, Speed selection and stability of wavefronts for delayed monostable reaction-diffusion equations, J. Dyn. Differ. Equations, 28 (2016), 1265-1292. doi: 10.1007/s10884-015-9482-6
|
| [64] |
Z. C. Wang, W. T. Li, S. Ruan, Traveling fronts in monostable equations with nonlocal delayed effects, J. Dyn. Differ. Equations, 20 (2008), 573-607. doi: 10.1007/s10884-008-9103-8
|
| [65] |
A. Solar, S. Trofimchuk, Asymptotic convergence to pushed wavefronts in a monostable equation with delayed reaction, Nonlinearity, 28 (2015), 2027-2052. doi: 10.1088/0951-7715/28/7/2027
|
| [66] | H. Berestycki, L. Nirenberg, Traveling waves in cylinders, Inst. H. Poincare Anal. Non. Lineaire, 9 (1992), 497-572. |
| [67] |
H. L. Smith, X. Q. Zhao, Global asymptotic stability of traveling waves in delayed reactiondiffusion equations, SIAM J. Math. Anal., 31 (2000), 514-534. doi: 10.1137/S0036141098346785
|
| [68] |
S. L. Wu, T. C. Niu, C. H. Hsu, Global asymptotic stability of pushed traveling fronts for monostable delayed reaction-diffusion equations, Discrete Contin. Dyn. Syst. A, 37 (2017), 3467-3486. doi: 10.3934/dcds.2017147
|
| [69] |
O. Diekmann, H. G. Kaper, On the bounded solutions of a nonlinear convolution equation, Nonlinear Anal., 2 (1978), 721-737. doi: 10.1016/0362-546X(78)90015-9
|
| [70] | E. Trofimchuk, M. Pinto, S. Trofimchuk, Monotone waves for non-monotone and non-local monostable reaction-diffusion equations, J. Differ. Equations, 261 (2016), 203-1236. |
| [71] |
O. Bonnefon, J. Garnier, F. Hamel, L. Roques, Inside dynamics of delayed traveling waves, Math. Model. Nat. Phenom., 8 (2013), 42-59. doi: 10.1051/mmnp/20138305
|
| [72] |
M. Jankovic, S. Petrovskii, Are time delays always destabilizing? Revisiting the role of time delays and the Allee effect, Theor. Ecol., 7 (2014), 335-349. doi: 10.1007/s12080-014-0222-z
|
| [73] | S. Ma, J. Wu, Existence, uniqueness and asymptotic stability of traveling wavefronts in a nonlocal delayed diffusion equation, J. Dyn. Differ. Equations, 19 (2007), 391-436. |
| [74] |
Z. C. Wang, W. T. Li, S. Ruan, Existence and stability of traveling wave fronts in reaction advection diffusion equations with nonlocal delay, J. Differ. Equations, 238 (2007), 153-200. doi: 10.1016/j.jde.2007.03.025
|
| [75] | A. Ducrot, M. Marion, V. Volpert, Spectrum of some integro-differential operators and stability of travelling waves, Nonlinear Anal. 74 (2011), 4455-4473. |
| [76] |
J. Wu, X. Zou, Traveling wave fronts of reaction-diffusion systems with delay, J. Dyn. Differ. Equations, 13 (2001), 651-687. doi: 10.1023/A:1016690424892
|
| [77] |
T. Faria, S. Trofimchuk, Non-monotone travelling waves in a single species reaction-diffusion equation with delay, J. Differ. Equations, 228 (2006), 357-376. doi: 10.1016/j.jde.2006.05.006
|
| [78] |
K. Hasík, J. Kopfová, P. Nábělková, S. Trofimchuk, Traveling waves in the nonlocal KPP-Fisher equation: different roles of the right and the left interactions, J. Differ. Equations, 260 (2016), 6130-6175. doi: 10.1016/j.jde.2015.12.035
|
| [79] |
K. Hasík, S. Trofimchuk, Slowly oscillating wavefronts of the KPP-Fisher delayed equation, Discrete Contin. Dyn. Syst. A, 34 (2014), 3511-3533. doi: 10.3934/dcds.2014.34.3511
|
| [80] | E. Trofimchuk, V. Tkachenko and S. Trofimchuk, Slowly oscillating wave solutions of a single species reaction-diffusion equation with delay, J. Differ. Equations, 245 (2008) 2307-2332. |
| [81] |
Z. Chladná, K. Hasík, J. Kopfova, P. Nábělková, S. Trofimchuk, Nonlinearly determined wavefronts of the Nicholson's diffusive equation: when small delays are not harmless, J. Differ. Equations, 268 (2020), 5156-5178. doi: 10.1016/j.jde.2019.11.007
|
| [82] | K. Hasík, J. Kopfova, P. Nábělková, S. Trofimchuk, On the geometric diversity of wavefronts for the scalar Kolmogorov ecological equation, J. Nonlinear Sci., 2020 (2020), 1-38. |
| [83] | E. Trofimchuk, M. Pinto, S. Trofimchuk, Existence and uniqueness of monotone wavefronts in a nonlocal resource-limited model, Proc. R. Soc. Edinburgh, Sect. A, 2020 (2020) 1-22. |
| [84] |
S. Ai, Traveling wave fronts for generalized Fisher equations with spatio-temporal delays, J. Differ. Equations, 232 (2007), 104-133. doi: 10.1016/j.jde.2006.08.015
|
| [85] | K. Hasík, J. Kopfova, P. Nábělková, S. Trofimchuk, On pushed wavefronts in monostable equation with unimodal delayed reaction, work in progress. |
| [86] | T. Yi, Y. Chen, J. Wu, Unimodal dynamical systems: Comparison principles, spreading speeds and travelling waves, J. Differ. Equations, 254 (2013), 3538-3572. |
| [87] |
C. Gomez, H. Prado, S. Trofimchuk, Separation dichotomy and wavefronts for a nonlinear convolution equation, J. Math. Anal. Appl., 420 (2014), 1-19. doi: 10.1016/j.jmaa.2014.05.064
|
| [88] |
S. Ma, Traveling waves for non-local delayed diffusion equations via auxiliary equations, J. Differ. Equations, 237 (2007), 259-277. doi: 10.1016/j.jde.2007.03.014
|
| [89] |
E. Trofimchuk, S. Trofimchuk, Admissible wavefront speeds for a single species reactiondiffusion equation with delay, Discrete Contin. Dyn. Syst. A, 20 (2008), 407-423. doi: 10.3934/dcds.2008.20.407
|
| [90] |
E. Trofimchuk, P. Alvarado, S. Trofimchuk, On the geometry of wave solutions of a delayed reaction-diffusion equation, J. Differ. Equations, 246 (2009), 1422-1444. doi: 10.1016/j.jde.2008.10.023
|
| [91] |
J. Fang, X. Q. Zhao, Monotone wavefronts of the nonlocal Fisher-KPP equation, Nonlinearity, 24 (2011), 3043-3054. doi: 10.1088/0951-7715/24/11/002
|
| [92] |
A. Gomez, S. Trofimchuk, Monotone traveling wavefronts of the KPP-Fisher delayed equation, J. Differ. Equations, 250 (2011), 1767-1787. doi: 10.1016/j.jde.2010.11.011
|
| [93] | J. K. Hale, X. B. Lin, Heteroclinic orbits for retarded functional differential equations, J. Differ. Equations, 65 (1985), 175-202. |
| [94] |
T. Faria, W. Huang, J. Wu, Traveling waves for delayed reaction-diffusion equations with nonlocal response, Proc. R. Soc. A, 462 (2006), 229-261. doi: 10.1098/rspa.2005.1554
|
| [95] |
W. Huang, Traveling waves connecting equilibrium and periodic orbit for reaction-diffusion equations with time delay and nonlocal response, J. Differ. Equations, 244 (2008), 1230-1254. doi: 10.1016/j.jde.2007.10.001
|
| [96] |
W. Huang, D. Duehring, Periodic travelling wave solutions for a reaction-diffusion equation with time delay and non-local response, J. Dyn. Differ. Equations, 19 (2007), 457-477. doi: 10.1007/s10884-006-9048-8
|
| [97] |
C. Ou, J. Wu, Persistence of wavefronts in delayed nonlocal reaction-diffusion equations, J. Differ. Equations, 235 (2007), 219-261. doi: 10.1016/j.jde.2006.12.010
|
| [98] |
C. Ou, J. Wu, Traveling wavefronts in a delayed food-limited population model, SIAM J. Math. Anal., 39 (2007), 103-125. doi: 10.1137/050638011
|
| [99] |
T. Faria, S. Trofimchuk, Positive traveling fronts for reaction-diffusion systems with distributed delay, Nonlinearity, 23 (2010), 2457-2481. doi: 10.1088/0951-7715/23/10/006
|
| [100] |
M. Aguerrea, S. Trofimchuk, G. Valenzuela, Uniqueness of fast traveling fronts in a single species reaction-diffusion equation with delay, Proc. R. Soc. Ser. A, 464 (2008), 2591-2608. doi: 10.1098/rspa.2008.0011
|
| [101] |
A. Gomez, S. Trofimchuk, Global continuation of monotone wavefronts, J. London Math. Soc., 89 (2014), 47-68. doi: 10.1112/jlms/jdt050
|
| [102] | V. Volpert, S. Trofimchuk, Global continuation of monotone waves for bistable delayed equations with unimodal nonlinearities, Nonlinearity, 32 2019, 2593-2632. |
| [103] | V. Volpert, Elliptic Partial Differential Equations. Volume 1. Fredholm Theory of Elliptic Problems in Unbounded Domains, Birkhäuser, (2011). |
| [104] | V. Volpert, Elliptic Partial Differential Equations. Volume 2. Reaction-Diffusion Equations, Birkhäuser, (2014). |
| [105] |
H. Berestycki, G. Nadin, B. Perthame, L. Ryzhik, The non-local Fisher-KPP equation: travelling waves and steady states, Nonlinearity, 22 (2009), 2813-2844. doi: 10.1088/0951-7715/22/12/002
|
| [106] |
S. Trofimchuk, V. Volpert, Traveling waves for a bistable reaction-diffusion equation with delay, SIAM J. Math. Anal., 50 (2018), 1175-1199. doi: 10.1137/17M1115587
|
| [107] |
S. Trofimchuk, V. Volpert, Existence of bistable waves for a nonlocal and nonmonotone reactiondiffusion equation, Proc. R. Soc. Edinburgh, Sect. A, 150 (2020), 721-739. doi: 10.1017/prm.2018.164
|
| [108] | G. Bocharov, A. Meyerhans, N. Bessonov, S. Trofimchuk, V. Volpert, Spatiotemporal dynamics of virus infection spreading in tissues, PLoS ONE, 11 (2016), e0168576. |
| [109] |
M. Alfaro, J. Coville, G. Raoul, Bistable travelling waves for nonlocal reaction diffusion equations, Discrete Contin. Dyn. Syst. A, 34 (2014), 1775-1791. doi: 10.3934/dcds.2014.34.1775
|
| [110] |
M. Alfaro, A. Ducrot, T. Giletti, Travelling waves for a non- monotone bistable equation with delay: existence and oscillations, Proc. London Math. Soc., 116 (2018), 729-759. doi: 10.1112/plms.12092
|
| [111] |
V. Volpert, Existence of waves for a bistable reaction-diffusion system with delay, J. Dyn. Differ. Equations, 32 (2020), 615-629. doi: 10.1007/s10884-019-09751-4
|
| [112] |
A. Solar, S. Trofimchuk, A simple approach to the wave uniqueness problem, J. Differ. Equations, 266 (2019), 6647-6660. doi: 10.1016/j.jde.2018.11.012
|
| [113] |
J. Coville, J.Dávila, S. Martínez, Nonlocal anisotropic dispersal with monostable nonlinearity, J. Differ. Equations, 244 (2008), 3080-3118. doi: 10.1016/j.jde.2007.11.002
|
| [114] |
G. Nadin, L. Rossi, L. Ryzhik, B. Perthame, Wave-like solutions for nonlocal reaction-diffusion equations: A toy model, Math. Model. Nat. Phenom., 8 (2013), 33-41. doi: 10.1051/mmnp/20138304
|
| [115] |
M. Mei, Ch. Ou, X. Q. Zhao, Global stability of monostable traveling waves for nonlocal timedelayed reaction-diffusion equations, SIAM J. Math. Anal., 42 (2010), 2762-2790. doi: 10.1137/090776342
|
| [116] | M. Mei, K. Zhang, Q. Zhang, Global stability of critical traveling waves with oscillations for time-delayed reaction-diffusion equation, Int. J. Numer. Anal. Model., 16 (2019), 375-397. |
| [117] |
A. Solar, Stability of semi-wavefronts for delayed reaction-diffusion equations, Nonlinear Differ. Equations Appl. NoDEA, 26 (2019), 33. doi: 10.1007/s00030-019-0580-8
|
| [118] |
A. Solar, Stability of non-monotone and backward waves for delay non-local reaction-diffusion equations, Discrete Contin. Dyn. Syst. A, 39 (2019), 5799-5823. doi: 10.3934/dcds.2019255
|
| [119] |
P. Ashwin, M. V. Bartuccelli, T. J. Bridges, S. A. Gourley, Travelling fronts for the KPP equation with spatio-temporal delay, Z. Angew. Math. Phys., 53 (2002), 103-122. doi: 10.1007/s00033-002-8145-8
|
| [120] |
M. K. Kwong, C. Ou, Existence and nonexistence of monotone traveling waves for the delayed Fisher equation, J. Differ. Equations, 249 (2010), 728-745. doi: 10.1016/j.jde.2010.04.017
|
| [121] | G. Bocharov, V. Volpert, B. Ludewig, A. Meyerhans, Mathematical Immunology of Virus Infections, Springer-Verlag, (2018). |
| [122] |
G. A. Bocharov, V. A. Volpert, A. L. Tasevich, Reaction-diffusion equations in Immunology, Comput. Math. Math. Phys., 58 (2018), 1967-1976. doi: 10.1134/S0965542518120059
|
| [123] | N. Bessonov, G. Bocharov, T. M. Touaoula, S. Trofimchuk, V. Volpert, Delay reaction-diffusion equation for infection dynamics, Discrete Contin. Dyn. Syst. B, 24 (2019) 2073-2091. |
| [124] | T. M. Touaoula, M. N. Frioui, N. Bessonov, V. Volpert, Dynamics of solutions of a reactiondiffusion equation with delayed inhibition, Discrete Contin. Dyn. Syst. S, 13 2020, 2425-2442. |
| [125] | N. Bessonov, G. Bocharov, V. Volpert, Space and genotype-dependent virus distribution during infection progression, work in progress. |
| [126] | N. Bessonov, G. Bocharov, A. Meyerhans, V. Popov, V. Volpert, Nonlocal reaction-diffusion model of viral evolution: emergence of virus strains, Mathematics, 8 2020,117. |
| [127] |
N. Bessonov, G. A. Bocharov, C. Leon, V. Popov, V. Volpert, Genotype-dependent virus distribution and competition of virus strains, Math. Mech. Compl. Syst., 8 (2020), 101-126. doi: 10.2140/memocs.2020.8.101
|
| [128] |
V. Volpert, Existence of reaction-diffusion waves in a model of immune response, Mediterr. J. Math., 17 (2020), 1-20. doi: 10.1007/s00009-019-1430-y
|
| [129] |
G. Bocharov, A. Meyerhans, N. Bessonov, S. Trofimchuk, V. Volpert, Modelling the dynamics of virus infection and immune response in space and time, Int. J. Parallel, Emergent Distrib. Syst., 34 (2019), 341-355. doi: 10.1080/17445760.2017.1363203
|
| [130] | V. Botella-Soler, M. Valderrama, B. Crépon, V. Navarro, M. Le Van Quyen, Large-scale cortical dynamics of sleep slow waves, PLoS ONE, 7 (2012), e30757. |
| [131] |
L. Muller, F. Chavane, J. Reynolds, T. J. Sejnowski, Cortical travelling waves: mechanisms and computational principles, Nat. Rev. Neurosci., 19 (2018), 255-268. doi: 10.1038/nrn.2018.20
|
| [132] | J. Y. Wu, X. Huang, C. Zhang, Propagating waves of activity in the neocortex: what they are, what they do, Neuroscientist, 14 (2008), 487-502. |
| [133] | H. R. Wilson, J. D. Cowan, A mathematical theory of the functional dynamics of cortical and thalamic nervous tissue, Kybernetik, 80 (1973), 55-80. |
| [134] |
H. Zhang, A. J. Watrous, A. Patel, J. Jacobs, Theta and alpha oscillations are travelling waves in the human neocortex, Neuron, 98 (2018), 1-13. doi: 10.1016/j.neuron.2018.03.027
|
| [135] | J. Senk, K. Korvasova, J. Schuecker, E. Hagen, T. Tetzlaff, M. Diesmann, et al., Conditions for travelling waves in spiking neural networks, Phys. Rev. Res., 2 (2020), 023174. |
| [136] |
D. J. Pinto, G. B. Ermentrout, Spatially structured activity in synapticalaly coupled neuronal networks: Ⅱ. Lateral inhibition and standing pulses, SIAM J. Appl. Math., 62 (2001), 226-243. doi: 10.1137/S0036139900346465
|
| [137] |
A. Beuter, A. Balossier, S. Trofimchuk, V. Volpert, Modeling of post-stroke stimulation of cortical tissue, Math. Biosci., 305 (2018), 146-159. doi: 10.1016/j.mbs.2018.08.014
|
| [138] | A. Moussaoui, V. Volpert, Speed of wave propagation for a nonlocal reaction-diffusion equation, Appl. Anal., 2018 (2018), 1-15. |
| [139] |
N. Bessonov, A. Beuter, S. Trofimchuk, V. Volpert, Dynamics of periodic waves in a neural field model, Mathematics, 8 (2020), 1076. doi: 10.3390/math8071076
|
| [140] |
F. M. Atay, A. Hutt, Neural Fields with Distributed Transmission Speeds and Long-Range Feedback Delays, SIAM J. Appl. Dyn. Syst., 5 (2006), 670-698. doi: 10.1137/050629367
|
| [141] |
N. A. Venkov, S. Coombes, P. C. Matthews, Dynamic instabilities in scalar neural field equations with space-dependent delays, Phys. D, 232 (2007), 1-15. doi: 10.1016/j.physd.2007.04.011
|
| [142] |
H. G. E. Meijer, S. Coombes, Travelling waves in a neural field model with refractoriness, J. Math. Biol., 68 (2014), 1249-1268. doi: 10.1007/s00285-013-0670-x
|
| [143] |
M. Adimy, A. Chekroun, B. Kazmierczak, Traveling waves in a coupled reaction-diffusion and difference model of hematopoiesis, J. Differ. Equations, 262 (2017), 4085-4128. doi: 10.1016/j.jde.2016.12.009
|
| [144] |
M. Jankovic, S. Petrovskii, M. Banerjee, Delay driven spatio-temporal chaos in single species population dynamics models, Theor. Popul. Biol., 110 (2016), 1-62. doi: 10.1016/j.tpb.2016.03.003
|
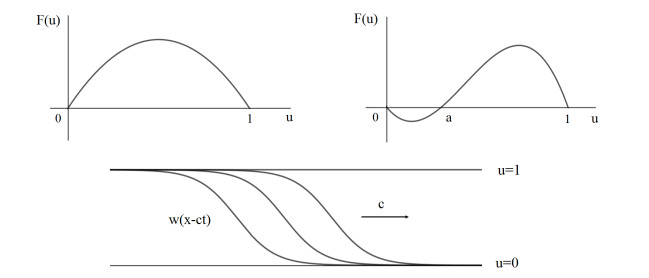
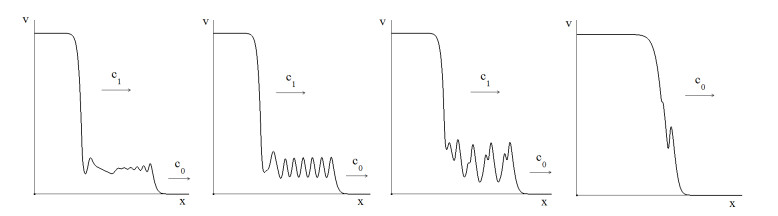
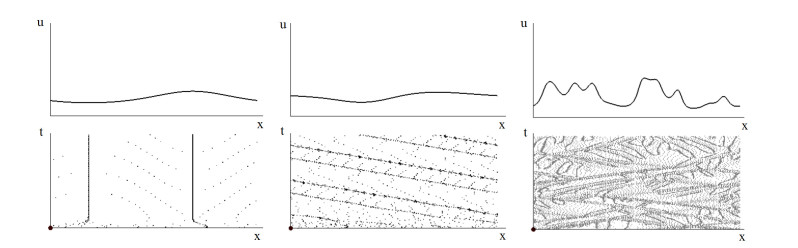
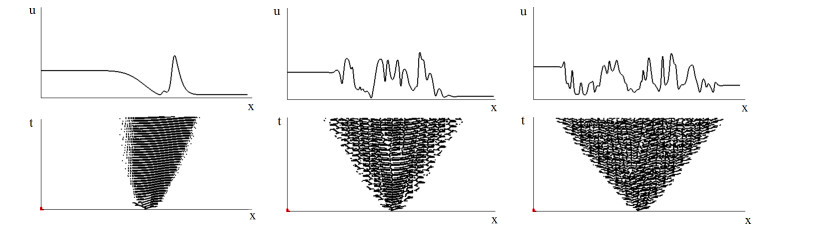
Figures(4)

Sergei Trofimchuk, Vitaly Volpert. Traveling waves in delayed reaction-diffusion equations in biology[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6487-6514. doi: 10.3934/mbe.2020339







 DownLoad:
DownLoad: