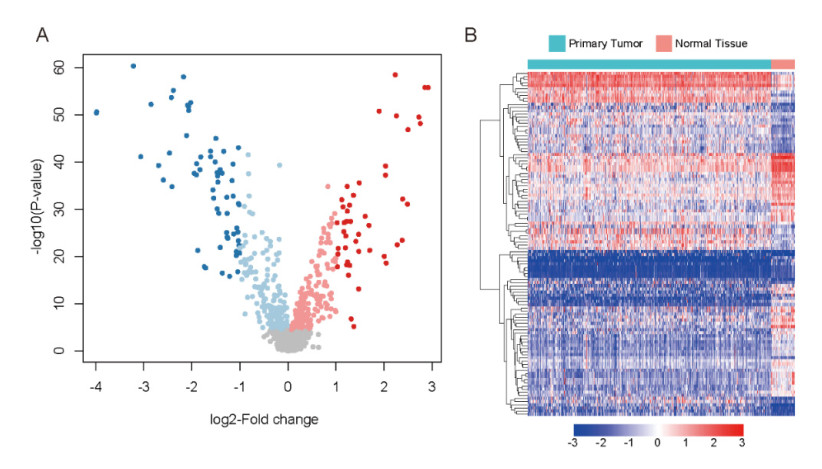
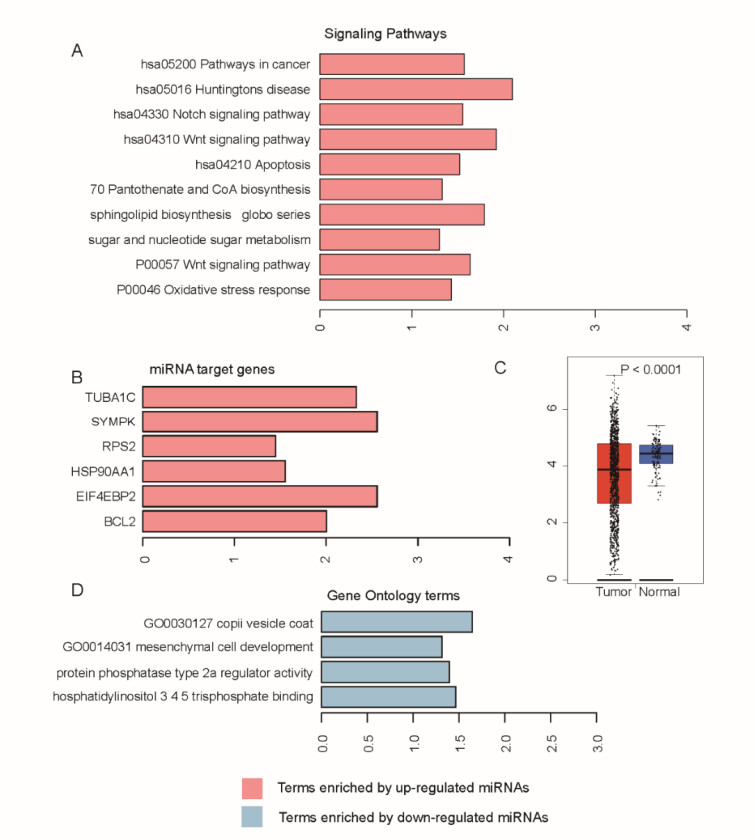
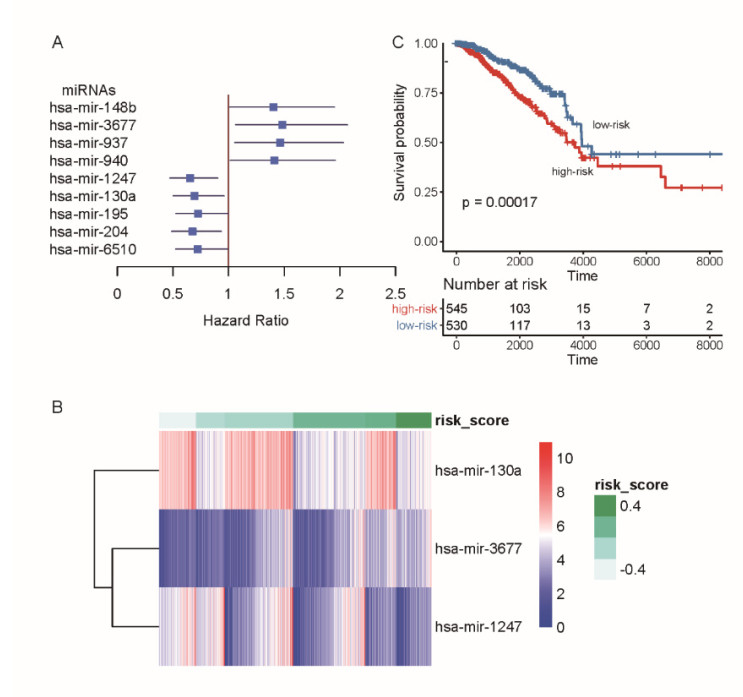
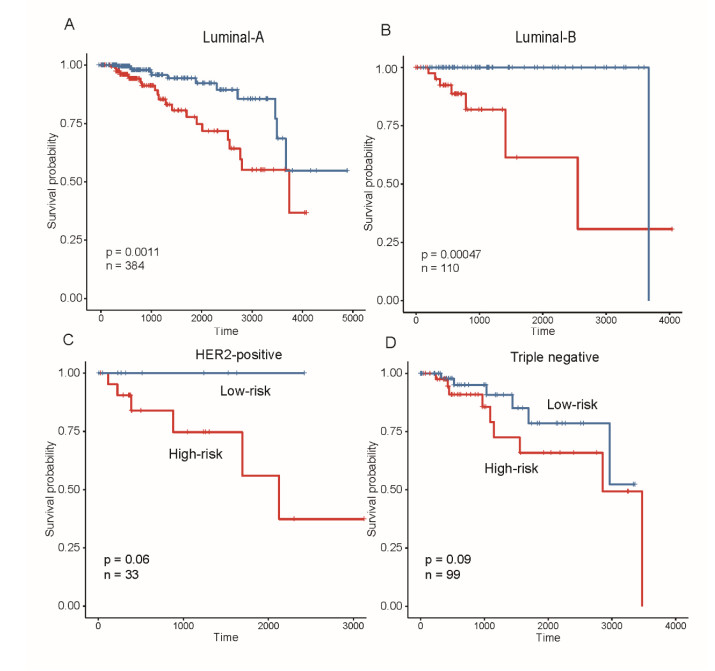
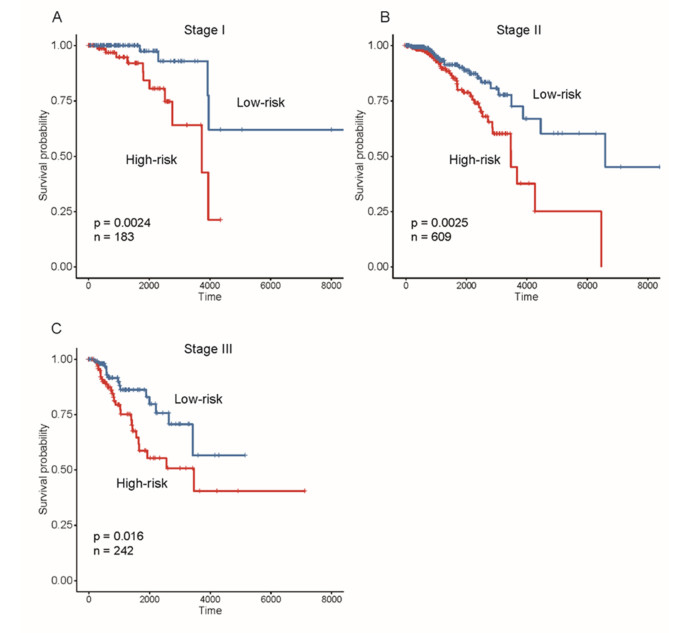
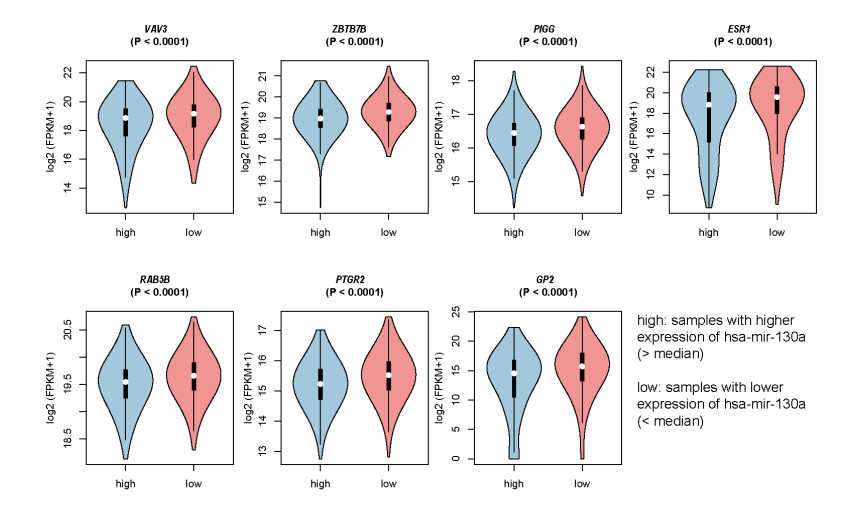
Breast cancer is a commonly diagnosed cancer in women, and one of the leading causes of cancer-related death among female patients However, the key microRNAs involved in its tumorigenesis and microRNAs of prognostic values have not been fully understood. In the present study, we aimed to perform a systematic analysis of microRNA expression profiles to identify some key microRNAs associated with tumor initiation and prognosis. Using TCGA breast cancer datasets, we identified 110 differentially expressed microRNAs. The functional enrichment analysis of the upregulated microRNAs revealed signaling transduction pathways, such as Notch and Wnt signaling pathway, and metabolism-related pathways such as sugar and nucleotide sugar metabolism, and oxidative stress response. Moreover, multivariable Cox model based on three variables of hsa-mir-130a, hsa-mir-3677, and hsa-mir-1247 stratified patients into high-risk and low-risk groups, which showed significant prognostic difference. In addition, we also tested the performance of this model in patient cohorts of any specific breast cancer subtypes or different TNM stages. The high performance in risk prediction was also observed in all of breast cancer subtypes and TNM stages. We also observed that there were highly possible interactions between hsa-mir-130a and seven target genes. Among these target genes, VAV3 and ESR1 were predicted as the target genes of hsa-mir-130a, suggesting that hsa-mir-130a may function by regulating the expression of VAV3 and ESR1 in breast cancer. In conclusion, the stratification based on the multivariable Cox model showed high performance in risk prediction. The dysregulated microRNAs and prognostic microRNAs greatly improved our understanding of the microRNA-related molecular mechanism underlying breast cancer.
Citation: Dongchen Lu, Wei Han, Kai Lu. Identification of key microRNAs involved in tumorigenesis and prognostic microRNAs in breast cancer[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 2923-2935. doi: 10.3934/mbe.2020164
Breast cancer is a commonly diagnosed cancer in women, and one of the leading causes of cancer-related death among female patients However, the key microRNAs involved in its tumorigenesis and microRNAs of prognostic values have not been fully understood. In the present study, we aimed to perform a systematic analysis of microRNA expression profiles to identify some key microRNAs associated with tumor initiation and prognosis. Using TCGA breast cancer datasets, we identified 110 differentially expressed microRNAs. The functional enrichment analysis of the upregulated microRNAs revealed signaling transduction pathways, such as Notch and Wnt signaling pathway, and metabolism-related pathways such as sugar and nucleotide sugar metabolism, and oxidative stress response. Moreover, multivariable Cox model based on three variables of hsa-mir-130a, hsa-mir-3677, and hsa-mir-1247 stratified patients into high-risk and low-risk groups, which showed significant prognostic difference. In addition, we also tested the performance of this model in patient cohorts of any specific breast cancer subtypes or different TNM stages. The high performance in risk prediction was also observed in all of breast cancer subtypes and TNM stages. We also observed that there were highly possible interactions between hsa-mir-130a and seven target genes. Among these target genes, VAV3 and ESR1 were predicted as the target genes of hsa-mir-130a, suggesting that hsa-mir-130a may function by regulating the expression of VAV3 and ESR1 in breast cancer. In conclusion, the stratification based on the multivariable Cox model showed high performance in risk prediction. The dysregulated microRNAs and prognostic microRNAs greatly improved our understanding of the microRNA-related molecular mechanism underlying breast cancer.

| [1] |
G. A. Colditz, K. Bohlke, Priorities for the primary prevention of breast cancer, CA Cancer J. Clin., 64 (2014), 186-194. doi: 10.3322/caac.21225

|
| [2] |
U. Veronesi, P. Boyle, A. Goldhirsch R. Orecchia, G. Viale, Breast cancer, Lancet, 365 (2005), 1727-1741. doi: 10.1016/S0140-6736(05)66546-4
|
| [3] | Collaborative Group on Hormonal Factors in Breast Cancer, Familial breast cancer: Collaborative reanalysis of individual data from 52 epidemiological studies including 58,209 women with breast cancer and 101,986 women without the disease, Lancet, 358 (2001), 1389-1399. |
| [4] |
F. J. Nassar, R. Nasr, R. Talhouk, MicroRNAs as biomarkers for early breast cancer diagnosis, prognosis and therapy prediction, Pharmacol. Ther., 172 (2017), 34-49. doi: 10.1016/j.pharmthera.2016.11.012
|
| [5] |
T. Sorlie, C. M. Perou, R. Tibshirani, T. Aas, S. Geisler S, H. Johnsen, et al., Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications, Proc. Natl. Acad. Sci. U. S. A., 98 (2001), 10869-10874. doi: 10.1073/pnas.191367098
|
| [6] | A. Prat, E. Pineda, B Adamo, P. Galvan, A. Fernandez, L. Gaba, Clinical implications of the intrinsic molecular subtypes of breast cancer, Breast, 24 (2015), S26-35. |
| [7] |
O. Yersal, S. Barutca, Biological subtypes of breast cancer: Prognostic and therapeutic implications, World J. Clin. Oncol., 5 (2014), 412-424. doi: 10.5306/wjco.v5.i3.412
|
| [8] |
J. N. Goh, S. Y. Loo, A. Datta, K. S. Siveen, W. N. Yap, W. Cai, et al., microRNAs in breast cancer: Regulatory roles governing the hallmarks of cancer, Biol. Rev., 91 (2016), 409-428. doi: 10.1111/brv.12176
|
| [9] |
D. P. Bartel, MicroRNAs: Target recognition and regulatory functions, Cell, 136 (2009), 215-233. doi: 10.1016/j.cell.2009.01.002
|
| [10] | V. A. Gennarino, M. Sardiello, R. Avellino, N. Meola, V. Maselli, S. Anand, et al., MicroRNA target prediction by expression analysis of host genes, Genome Res., 19 (2009), 481-490. |
| [11] |
A. Esquela-Kerscher, F. J. Slack, Oncomirs - microRNAs with a role in cancer, Nat. Rev. Cancer, 6 (2006), 259-269. doi: 10.1038/nrc1840
|
| [12] | M. Goldman, B. Craft, A. Brooks, J. Zhu, D. Haussler, The UCSC Xena Platform for cancer genomics data visualization and interpretation, BioRxiv, 2018 (2018), 326470. |
| [13] |
C. Backes, Q. T. Khaleeq, E. Meese, A. Keller, miEAA: MicroRNA enrichment analysis and annotation, Nucleic Acids Res., 44 (2016), W110-W116. doi: 10.1093/nar/gkw345
|
| [14] |
C. H. Chou, S. Shrestha, C. D. Yang, N. W. Chang, Y. L. Lin, K. W. Liao, et al., miRTarBase update 2018: a resource for experimentally validated microRNA-target interactions, Nucleic Acids Res., 46 (2018), D296-D302. doi: 10.1093/nar/gkx1067
|
| [15] | L. E. Brown, I. Tsamardinos, C. F. Aliferis, A novel algorithm for scalable and accurate Bayesian network learning, Studies in health technology and informatics, 107 (2004), 711-715. |
| [16] |
M. Q. Cao, A. B. You, X. D. Zhu, W. Zhang, Y. Y. Zhang, S. Z. Zhang, et al., miR-182-5p promotes hepatocellular carcinoma progression by repressing FOXO3a, J. Hematol. Oncol., 11 (2018), 12. doi: 10.1186/s13045-018-0555-y
|
| [17] | X. Fei, J. Zhang, Y. Zhao, M. Sun, H. Zhao, S. Li, miR-96 promotes invasion and metastasis by targeting GPC3 in non-small cell lung cancer cells, Oncol. Lett., 15 (2018), 9081-9086. |
| [18] |
Y. H. Feng, C. J. Tsao, Emerging role of microRNA-21 in cancer, Biomed. Rep., 5 (2016), 395-402. doi: 10.3892/br.2016.747
|
| [19] |
Q. Fu, Y. Du, C. Yang, D. Zhang, N. Zhang, X. Liu, et al., An oncogenic role of miR-592 in tumorigenesis of human colorectal cancer by targeting Forkhead Box O3A (FoxO3A), Expert Opin. Ther. Targets, 20 (2016), 771-782. doi: 10.1080/14728222.2016.1181753
|
| [20] | J. M. Gao, L. Z. Huang, Z. G. Huang, R. Q. He, Clinical value and potential pathways of miR-183-5p in bladder cancer: A study based on miRNA-seq data and bioinformatics analysis, Oncol. Lett., 15 (2018), 5056-5070. |
| [21] |
D. Li, X. Liu, L. Lin, J. Hou, N. Li, C. Wang, et al., MicroRNA-99a inhibits hepatocellular carcinoma growth and correlates with prognosis of patients with hepatocellular carcinoma, J. Biol. Chem., 286 (2011), 36677-36685. doi: 10.1074/jbc.M111.270561
|
| [22] |
Z. Li, H. Lei, M. Luo, Y. Wang, L. Dong, Y. Ma, et al., DNA methylation downregulated mir-10b acts as a tumor suppressor in gastric cancer, Gastric Cancer, 18 (2015), 43-54. doi: 10.1007/s10120-014-0340-8
|
| [23] |
M. Pajic, D. Froio, S. Daly, L. Doculara, E. Millar, P. H. Graham, et al., miR-139-5p Modulates Radiotherapy Resistance in Breast Cancer by Repressing Multiple Gene Networks of DNA Repair and ROS Defense, Cancer Res., 78 (2018), 501-515. doi: 10.1158/0008-5472.CAN-16-3105
|
| [24] | F. Peng, T. T. Li, K. L. Wang, G. Q. Xiao, J. H. Wang, H. D. Zhao, et al., H19/let-7/LIN28 reciprocal negative regulatory circuit promotes breast cancer stem cell maintenance, Cell Death Dis., 8 (2017), e2569. |
| [25] |
H. Wang, C. Hang, X. L. Ou, J. S. Nie, Y. T. Ding, S. G. Xue, et al., MiR-145 functions as a tumor suppressor via regulating angiopoietin-2 in pancreatic cancer cells, Cancer Cell Int., 16 (2016), 65. doi: 10.1186/s12935-016-0331-4
|
| [26] |
B. M. Krishna, S. Jana, J. Singhal, D. Horne, S. Awasthi, R. Salgia, et al., Notch signaling in breast cancer: From pathway analysis to therapy, Cancer Lett., 461 (2019), 123-131. doi: 10.1016/j.canlet.2019.07.012
|
| [27] |
P. Yin, W. Wang, Z. Zhang, Y. Bai, J. Gao, C. Zhao, Wnt signaling in human and mouse breast cancer: Focusing on Wnt ligands, receptors and antagonists, Cancer Sci., 109 (2018), 3368-3375. doi: 10.1111/cas.13771
|
| [28] |
S. J. Dawson, N. Makretsov, F. M. Blows, K. E. Driver, E. Provenzano, J. Le Quesne, et al., BCL2 in breast cancer: a favourable prognostic marker across molecular subtypes and independent of adjuvant therapy received, Br. J. Cancer, 103 (2010), 668-675. doi: 10.1038/sj.bjc.6605736
|
| [29] |
H. Aguilar, A. Urruticoechea, P. Halonen, K. Kiyotani, T. Mushiroda, X. Barril, et al., VAV3 mediates resistance to breast cancer endocrine therapy, Breast Cancer Res., 16 (2014), R53. doi: 10.1186/bcr3664
|
| [30] |
C. S. Ross-Innes, R. Stark, K. A. Holmes, D. Schmidt, C. Spyrou, R. Russell, et al., Cooperative interaction between retinoic acid receptor-alpha and estrogen receptor in breast cancer, Genes Dev., 24 (2010), 171-182. doi: 10.1101/gad.552910
|
| [31] | L. X. Yan, X. F. Huang, Q. Shao, M. Y. Huang, L. Deng, Q. L. Wu, et al., MicroRNA miR-21 overexpression in human breast cancer is associated with advanced clinical stage, lymph node metastasis and patient poor prognosis, RNA, 14 (2018), 2348-2360. |
| [32] |
P. Li, C. Sheng, L. Huang, H. Zhang, L. Huang, Z. Cheng, et al., MiR-183/-96/-182 cluster is up-regulated in most breast cancers and increases cell proliferation and migration, Breast Cancer Res., 16 (2014), 473. doi: 10.1186/s13058-014-0473-z
|
Figures(6) / Tables(1)

Dongchen Lu, Wei Han, Kai Lu. Identification of key microRNAs involved in tumorigenesis and prognostic microRNAs in breast cancer[J]. Mathematical Biosciences and Engineering, 2020, 17(4): 2923-2935. doi: 10.3934/mbe.2020164







 DownLoad:
DownLoad: