Citation: Shunyi Li. Hopf bifurcation, stability switches and chaos in a prey-predator system with three stage structure and two time delays[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6934-6961. doi: 10.3934/mbe.2019348

| [1] | W. Aiello and H. Freedman, A time-delay model of single-species growth with stage structure, Math. Biosci., 101 (1990), 139–153. |
| [2] | W. Aiello, H. Freedman and J. Wu, Analysis of a model representing stage-structured populations growth with stage-dependent time delay, SIAM J. Appl. Math., 3 (1992), 855–869. |
| [3] | W. Wang and L. Chen, A predator-prey system with stage-structure for predator, Comp. Math. Appl., 33 (1997), 83–91. |
| [4] | S. Liu, L. Chen and R. Agarwal, Recent progress on stage-structured population dynamics, Math. Comput. Model., 36 (2002), 1319–1360. |
| [5] | S. Gao, Models for single species with three life history stages and cannibalism, J. Biomath., 20 (2005), 385–391. |
| [6] | S. Yang and B. Shi, Periodic solution for a three-stage-structured predator-prey system with time delay, J. Math. Anal. Appl., 341 (2008), 287–294. |
| [7] | H. Smith, An Introduction to Delay Differential Equations with Applications to the Life Sciences, Springer Science+Business Media, LLC, 201l. |
| [8] | E. Beretta and D. Breda, Discrete or distributed delay? Effects on stability of population growth, Math. Biosci. Eng., 13 (2016), 19–41. |
| [9] | Z. Shen and J. Wei, Hopf bifurcation analysis in a diffusive predator-prey system with delay and surplus killing effect, Math. Biosci. Eng., 15 (2018), 693–715. |
| [10] | S. Li and Z. Xiong, Bifurcation analysis of a predator-prey system with sex-structure and sexual favoritism, Adv. Differ. Equ., 219 (2013), 1–24. |
| [11] | Z. Ma and S. Wang, A delay-induced predator Cprey model with Holling type functional response and habitat complexity, Nonl. Dyna., 93 (2018), 1519–1544. |
| [12] | S. Kundu and S. Maitra Dynamical behaviour of a delayed three species predator Cprey model with cooperation among the prey species, Nonl. Dyna., 92 (2018), 627–643. |
| [13] | L. Li and J. Shen, Bifurcations and Dynamics of a Predator CPrey Model with Double Allee Effects and Time Delays, Int. J. Bifurc. Chaos, 28 (2018), 1–14. (No. 1850135) |
| [14] | T. Caraballo, R. Colucci and L. Guerrini, On a predator prey model with nonlinear harvesting and distributed delay, Comm. on Pure Appl. Anal., 17 (2018), 2703–2727. |
| [15] | X. Xu, Y. Wang and Y. Wang, Local bifurcation of a Ronsenzwing-MacArthur predator prey model with two prey-taxis, Math. Biosci. Eng., 16 (2019), 1786-1797. |
| [16] | S. Li, Y. Xue and W. Liu, Hopf bifurcation and global periodic solutions for a three-stage-structured prey-predator system with delays, Int. J. Info. Syst. Scie., 8 (2012), 142–156. |
| [17] | S. Li and X. Xue, Hopf bifurcation in a three-stage-structured prey-predator system with predator density dependent, Comm. Comp. Info. Scie., 288 (2012), 740–747. |
| [18] | S. Li and W. Liu, Global hopf bifurcation in a delayed three-stage-structured prey-predator system, Proceedings-5th Int. Conf. Info. Comp. Scie., (2012), 206–209. |
| [19] | J. Wu, Symmetric functional differential equations and neural networks with memory, Trans. Am. Math. Soc., 350 (1998), 4799–4838. |
| [20] | Z. Wang, A very simple criterion for characterizing the crossing direction of time-delay systems with delay-dependent parameters, Int. J. Bifu. Chaos, 22 (2012), 1–7. |
| [21] | J. Hale, Theory of Functional Differential Equations, Springer, New York, 1977. |
| [22] | D. Breda, S. Maset and R. Vermiglio, TRACE-DDE: a tool for robust analysis and characteristic equations for delay differential equations, Lect. Notes Cont. Info. Scie., 388 (2009), 145–155. |
| [23] | D. Breda, S. Maset and R. Vermiglio, Pseudospectral differencing methods for characteristic roots of delay differential equations, SIAM J. Sci. Comput., 27 (2005), 482–495. |
| [24] | D. Breda, S. Maset and R. Vermiglio, An adaptive algorithm for efficient computation of level curves of surfaces, Numer. Algorithms, 52 (2009), 605–628. |
| [25] | Y. Zhao, X. Yu and L. Wang, Bifurcation and control in an inertial two-neuron system with time delays, Int. J. Bifurc. Chaos, 22 (2012), 1–15. |
| [26] | S. Li, W. Liu and X. Xue, Hopf bifurcation, chaos and impulsive control in a sex-structured prey-predator system with time delay, J. Biomath., 30 (2015), 443–452. |
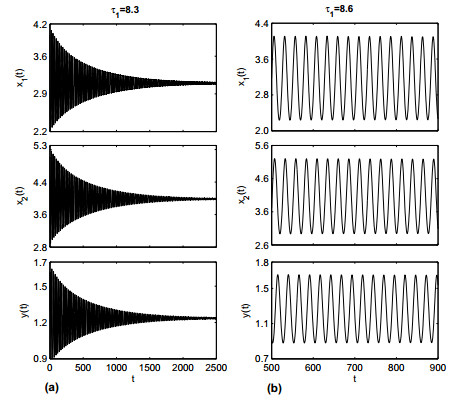
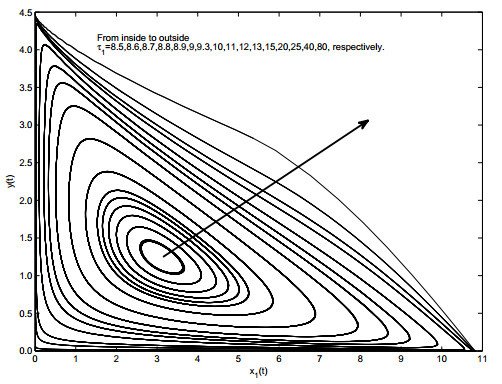
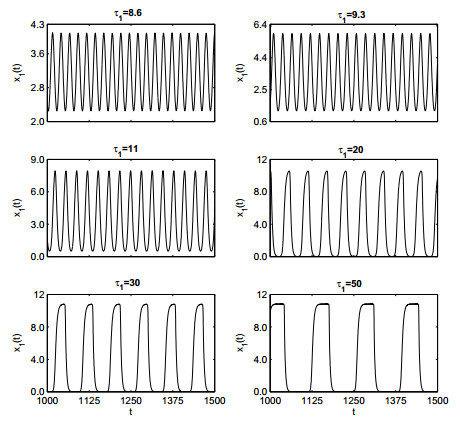
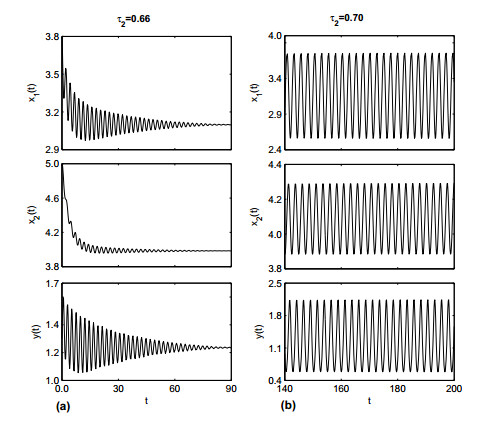
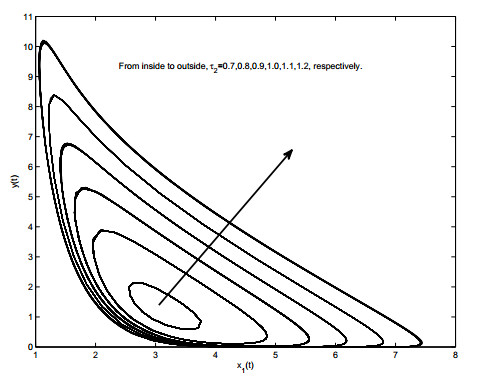
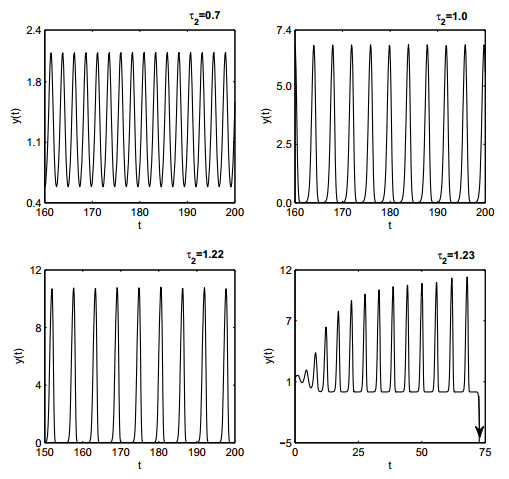
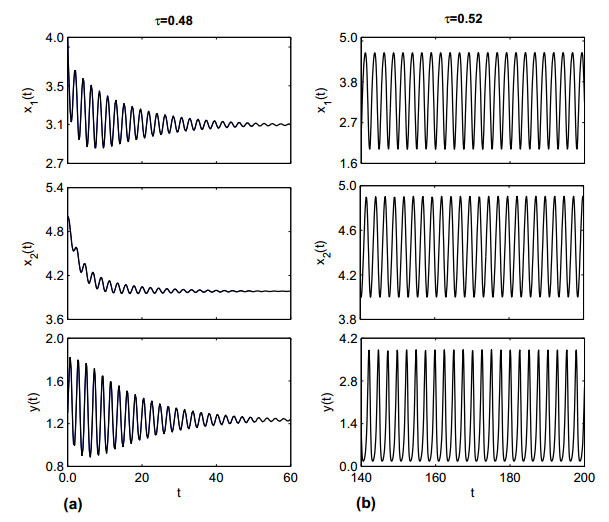
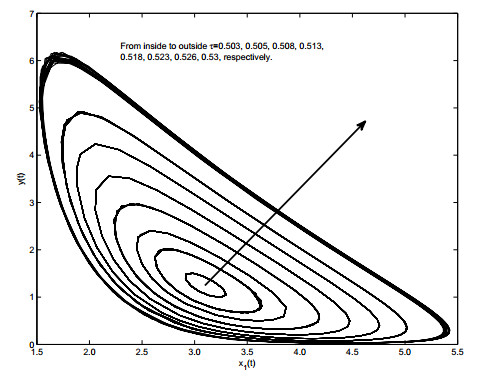
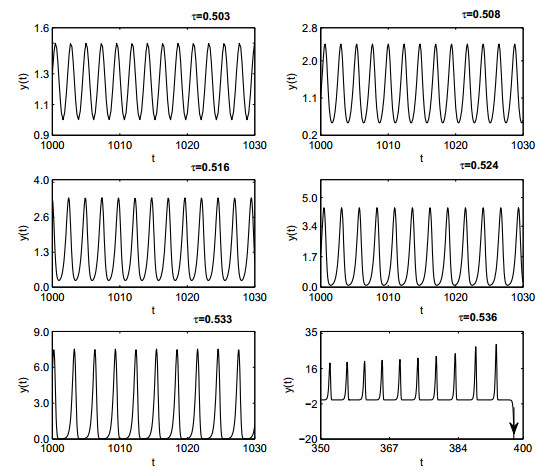
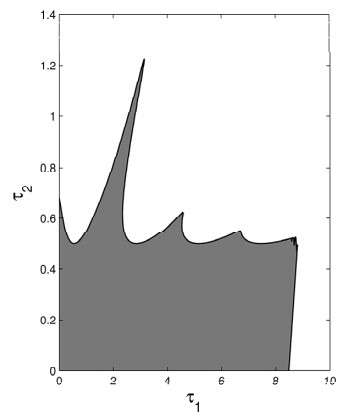
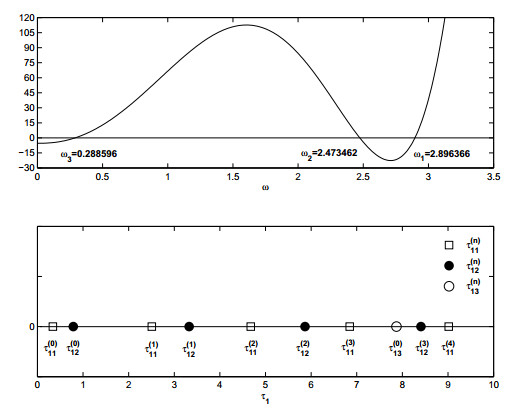
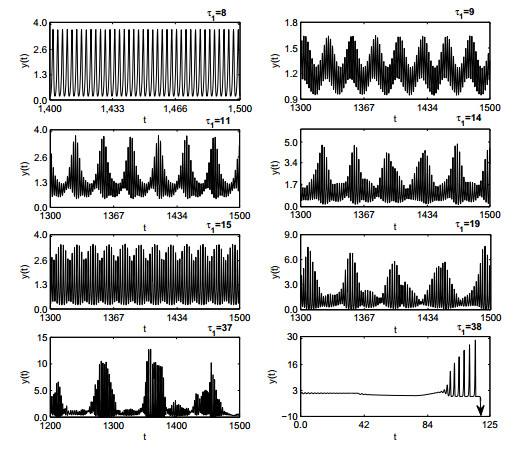
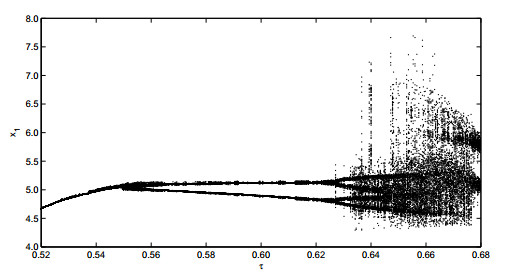
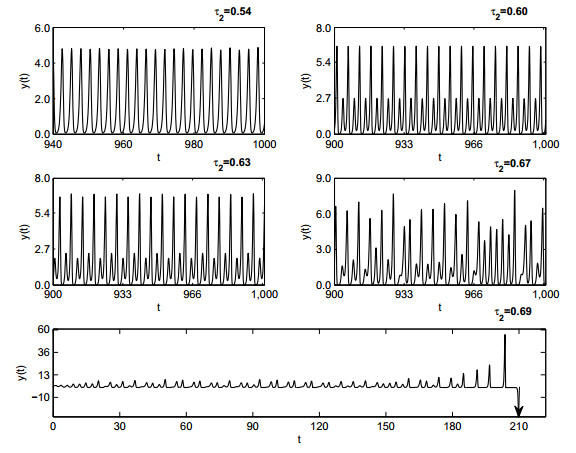
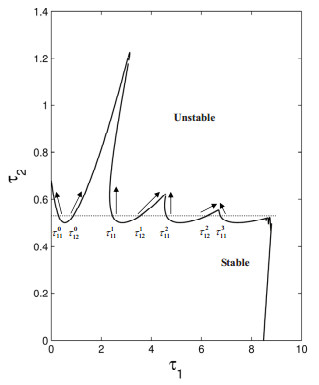
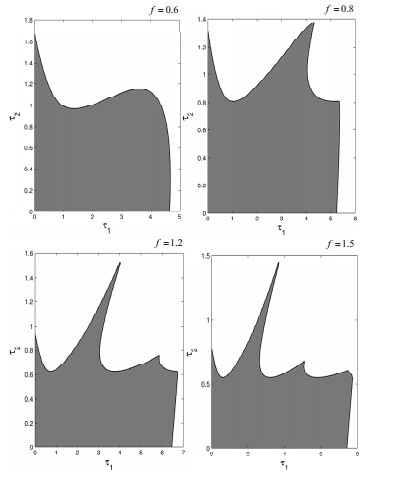
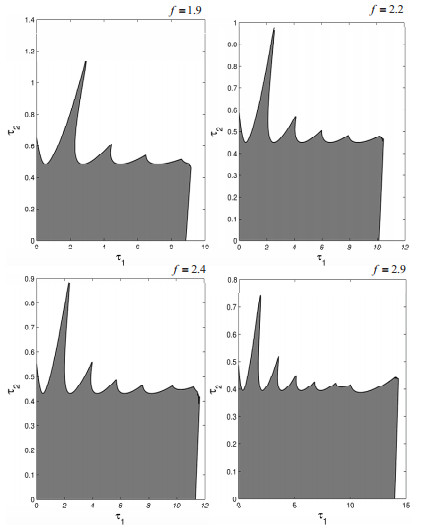
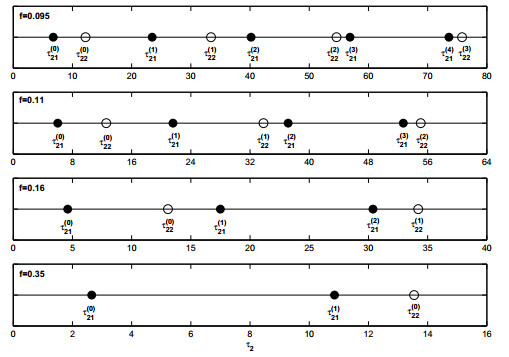
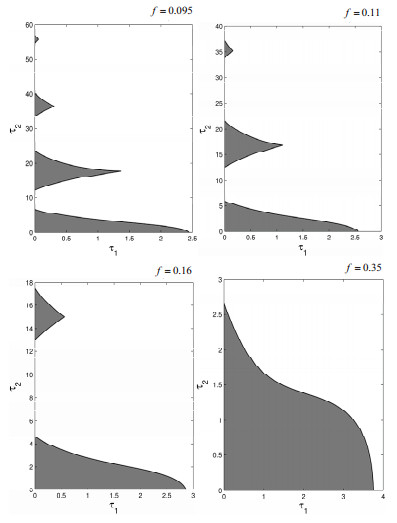
Figures(19)

Shunyi Li. Hopf bifurcation, stability switches and chaos in a prey-predator system with three stage structure and two time delays[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6934-6961. doi: 10.3934/mbe.2019348







 DownLoad:
DownLoad: